1. 引言
马氏珠母贝(Pinctada martensii Dunker)又称合浦珠母贝,是目前我国人工海水珍珠育珠的主要珍珠贝,海水珍珠养殖已成为广东、广西和海南沿海部分地区的支柱产业之一。我国自1965年成功地开展马氏珠母贝的人工育苗以来,在单一环境因子及多因子对马氏珠母贝人工育苗方面有不少研究[1] -[7] 。在多因子对受精、孵化及稚贝生长存活方面,喻达辉等[8] -[10] 研究了海水中离子对马氏珠母贝精子激活机制和离子选择性并对马氏珠母贝精子的实验生物学进行研究;沈亦平等[11] 对马氏珠母贝配子的发生和受精进行了细胞学观察;朱晓闻[12] 研究了温度、盐度、pH对马氏珠母贝稚贝生理学特征的联合效应;邓云翠[13] 等研究了Cd2+与Zn2+的联合效应对马氏珠母贝受精率的影响;王辉等[14] 研究了温度和盐度对马氏珠母贝精子激活率及活力的影响,但未对其受精和孵化的影响以及各环境因素之间是否存在互作进行进一步研究。
贝类生活与周围环境也密切相关,环境因素的优劣,能加速阻碍贝类的生长和存亡。如水温的高低不但影响了贝类幼虫的新陈代谢的强弱,还影响了饵料生物的繁殖与生长,也影响了酸碱度的变化等,这就间接地影响了贝类的生长[15] -[26] 。盐度的高低则直接影响成贝的生长和幼体发育[25] -[35] 。盐度不适导致幼虫死亡的主要原因是海水渗透压的改变超出了自身调节能力所致,贝类体内的组织和细胞要消耗大量的能量来调节渗透压。渗透压的改变不仅会降低动物的代谢速率,同时也会影响代谢过程的效率。
本研究通过二次回归组合正交设计,考察在不同温度和盐度下马氏珠母贝人工受精和胚胎发育的情况,并建立起对应的数学模型,研究结果将从理论上更好的指导马氏珠母贝的人工育苗。
2. 材料与方法
2.1. 实验材料
生物学材料:本试验所采用的亲贝来源于雷州市流沙镇,为同一批亲贝,在亲贝池中暂养,性腺饱满,活力较好。
实验设备:恒温加热棒、气泵、显微镜、血球计数板、胶头滴管、比重计、温度计、大烧杯、100 L白桶、海盐等。
2.2. 实验方法
2.2.1. 实验方案设计
本试验拟设定两个因子:温度和盐度,并采用二次回归组合正交设计考察对马氏贝受精率和孵化率的影响。二次回归组合正交设计方案如表1~表2。
2.2.2. 实验准备
按照所设计的梯度组合,分别在22只100 L白桶中配置好相应温度和盐度的海水,投入气石充气备用。以调整好温度和盐度的海水来配置人工受精过程中活化精卵所用的氨海水,浓度为万分之1.5,现配现用。实验分22个组,每组分别挑选性腺饱满的成熟雌贝2个和雄贝1个,备用。
2.2.3. 实验步骤
1) 解剖:解剖雌贝,将分离出来的性腺用双层纱布包裹住,放入盛有500 ml配置好温度盐度的氨海水的大烧杯中将性腺剪碎,滤出的卵液与氨海水混合均匀。取样计数后继续活化3~5 min。其中取卵以血球计数板计算单位体积卵数,再通过密度体积关系求出烧杯中的总卵数,取3次求平均值。
2) 活化:在活化卵子的同时解剖雄贝,剪破其性腺,有乳白色精液流出,用胶头滴管吸取少量精液置于250 ml配置好温度和盐度的氨海水的大烧杯中,混合均匀,活化3~5 min。
3) 受精:将活化后的精卵倒入1 L的大烧杯中,搅拌均匀,充分受精。整个受精过程中大烧杯放入相应温度的白桶中水浴,以保证整个人工受精都在实验所需的温度下进行。每30~40 min换水洗卵一次。受精一段时间后,受精卵下沉,在水体中形成明显的分层。不成熟的未受精卵和多余的精子悬浮在上层(镜检发现,虽然卵子在氨水的刺激下大部分变为圆形,但未出现受精的标志:第一极体。而下层的卵子大

Table 1. Level of factors
表1. 因素水平表
Table 2. Combination of factors level
表2. 试验因子水平组合表
部分受精,出现第一极体),用吸管小心地将上层的液体虹吸出,再加入同一温度和盐度的海水,静置。三次洗卵后可将大部分未受精卵、多余精子和氨水洗去,以保证孵化过程环境适宜。洗卵结束后分别取样镜检计数,通过密度体积关系求出受精卵的总量,取3次求平均值,再通过公式计算出各组合中马氏珠母贝精卵的受精率。受精率的计算公式为:
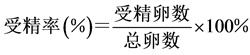
4) 洗卵:洗卵结束后,分别将各组受精卵倒入100 L的白桶中,控制好温度和盐度,充气使水面呈微波状,24 h后取样计数。在适宜温度和盐度(26℃, 29‰)的情况下,马氏珠母贝受精卵发育到D型幼虫需要19~24 h,本实验以24 h为标准。取样计数方法为:先将充气量加大,并搅动水体,待水体中幼虫趋于均匀分布后,用小烧杯在白桶中不同部位取样3次,每次量取100 ml海水,滴入少量甲醛溶液,轻轻搅拌均匀以固定和杀死D型幼虫。静置3~5 h后可观察到幼虫沉淀到烧杯的底部,小心吸去上清液,用滴管吸取底部沉淀在显微镜下计数,再根据公式求出孵化率。孵化率的计算公式为:
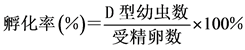
2.2.4. 数据处理与统计分析
试验数据由SAS8.2中的调用GLM模块处理。显著性水平设为a = 0.05。
3.结果
3.1. 温度和盐度对受精率的影响
试验分析结果如下。
根据表3可以得到数学模型的具体形式。
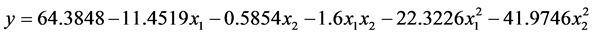
其中y为马氏珠母贝受精率的理论估计值,a为各项的偏回归系数。
失拟即回归模型选择不当造成的误差,进行失拟检验可验证回归模型选择是否恰当[36] 。根据表4可知,由于失拟F < F-crit,是不显著的(P > 0.05),说明回归模型的拟合度较高。通过表5可知,由于表中F > F-crit,说明回归方程在 水平上显著。
水平上显著。
得到总的回归方程后,根据表6对各项回归系数的检验可知,x1、x2与x1x2的回归系数的F均小于F-crit,是不显著的(P > 0.05);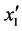 的F大于6.6079但小于16.2582,说明其在a = 0.05水平上显著(P < 0.05);
的F大于6.6079但小于16.2582,说明其在a = 0.05水平上显著(P < 0.05);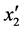 的F > F0.01,是极显著的(P < 0.01)。将各不显著项剔除,可以得到温度和盐度影响马氏珠母贝受精率的数学模型最终形式
的F > F0.01,是极显著的(P < 0.01)。将各不显著项剔除,可以得到温度和盐度影响马氏珠母贝受精率的数学模型最终形式

3.2. 温度和盐度对孵化率的影响
试验分析结果如下。
根据表7可以得到数学模型的具体形式
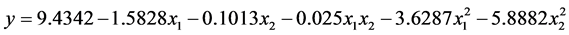
其中y为马氏珠母贝受精卵孵化率的理论估计值,a为各项的偏回归系数。
失拟即回归模型选择不当造成的误差,进行失拟检验可验证回归模型选择是否恰当。根据表8可
Table 3. Calculation of orthogonal combination quadratic regression
表3. 二次回归组合正交计算
注:表中Bj表示自变量与异变量乘积和,dj表示自变量平方和,bj表示偏回归系数,Qj表示偏回归平方和,下同。
Table 4. Analysis of variance for lack of fit test of model
表4. 模型失拟检验方差分析
注:F-crit表示临界值,分别为5%和1%显著水平,下同。
Table 5. Analysis of variance for regression model
表5. 回归模型方差分析
*表示差异显著(p < 0.05),**表示差异极显著(P < 0.01),下同。
知,由于失拟F < F-crit,是不显著的(P > 0.05),说明回归模型的拟合度较高。通过表9可知,由于表中F > F-crit,说明回归方程在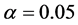 水平上显著。
水平上显著。
得到总的回归方程后,根据表10对各项回归系数的检验可知,x1、x2与x1x2的回归系数的F均小于
Table 6. Test of regression coefficient
表6. 回归系数检验
Table 7. Calculation of orthogonal combination quadratic regression
表7. 二次回归组合正交计算
Table 8. Analysis of variance for lack of fit test of model
表8. 模型失拟检验方差分析
F-crit,是不显著的(P > 0.05);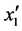 的F大于6.6079小于16.2582,说明其在a = 0.05水平上显著(P < 0.05);x2'的F > F0.01,是极显著的(P < 0.01)。将各不显著项剔除,可以得到温度和盐度影响马氏珠母贝受精卵孵化率的数学模型最终形式
的F大于6.6079小于16.2582,说明其在a = 0.05水平上显著(P < 0.05);x2'的F > F0.01,是极显著的(P < 0.01)。将各不显著项剔除,可以得到温度和盐度影响马氏珠母贝受精卵孵化率的数学模型最终形式
Table 9. Analysis of variance for regression model
表9. 回归模型方差分析
Table 10. Test of regression coefficient
表10. 回归系数检验
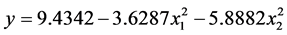
4. 讨论
生物对环境因素的变化是有一定范围,假如接近或超过这个范围的界限,这些因素就成了限制生物生活的因素。各种因素应当保持在生物生活的适当范围内,如果某种因素超过了生物的适应界限,这个因素便成了生物生存的决定性因素。目前看来,从单因素试验到多因素试验的研究是必然的趋势,而多因素试验处理组合数目随着因素数及每个因素的水平数增加而成指数增长。正交回归设计可以很好的处理多因素试验中的问题,它利用数理统计学观点,应用正交性原理,对多因素同时进行考察,既保证试验提供丰富的信息,又使试验数不过于庞大。关于马氏珠母贝的生长温度、盐度和饵料的研究已有报道,但对各因子之间的内在关系的研究较少。本研究通过二次回归组合正交设计,考察了在不同温度和盐度下马氏珠母贝人工受精和胚胎发育的情况,并建立起了对应的数学模型。
通常,贝类在最适温度下精卵受精与受精卵发育的速度最快,随着温度向高温和低温两端的迁移受精及孵化的时间相应会延长[37] 。马氏珠母贝在26℃时由受精卵发育到D型幼虫最快仅要19 h左右,随着温度的升高和降低,发育到D型幼虫的时间可延长到30~40 h以上[6] 。本试验分析的是24 h孵化率,由于对各组采样计数的时间一致,均为24 h取样计数,考虑到高温组和低温组受精和发育的时间延长,故实际受精率和孵化率会有一定的提高。在试验中发现,较低温度下的卵子受精率比较低,而在胚胎发育过程中,在适温范围以上时,出现畸形发育或很快死亡;在适温范围以下时,出现停止进一步发育的现象。通常在适温范围以内,即使相差几度,发育速度也有显著的差异,而且在发育初期对温度的适应范围较窄,随着发育的进行,适应范围愈来愈广[37] 。在较低和较高盐度下,卵子大部分不能受精,极少数的受精卵发育也不正常,出现如细胞分裂畸形,卵膜皱缩,卵膜破裂等情况,主要是因为精卵和胚胎调节渗透压的机制还不健全。通过对不同温度和盐度下马氏珠母贝受精率与孵化率的回归分析可以发现,在受精和胚胎发育过程中,温度和盐度之间的互作效应并不明显,未达到显著水平(P > 0.05)。从两个因子的回归系数检验来看,温度对受精和孵化的影响要略大于盐度。王辉[14] 也指出,温度的一次效应对马氏珠母贝精子的激活和活力极显著,温度和盐度的互作效应对精子激活率和活力的影响不显著,这与本实验的结果是相吻合的。
在马氏珠母贝贝苗的实际生产过程中,还有温度、盐度、酸碱度、溶氧、饵料和光照等环境因子影响着育苗效果。除温度和盐度以外的其他环境因子对马氏珠母贝受精和孵化的影响也有待进一步研究。

NOTES
*通讯作者。