1. 引言
乙酰辅酶A羧化酶(acetyl-CoA carboxylase, ACCase),是植物中脂肪酸从头合成途径中的限速酶,在质体内参与以乙酰辅酶A为前体生成丙二酰辅酶A的过程。ACCase由三部分组成,即生物素羧基载体蛋白(biotin carboxyl carrier protein, BCCP)、生物素羧化酶(biotin carboxylase, BC)和羧基转移酶(carboxyltransferase, CT)。由于ACCase对植物的正常生长发育具有关键作用,因此,农药公司以ACCase为靶标,开发研制了一系列除草剂,在市场推广至今。
2. ACCase类型和定位
2.1. ACCase的种类
在大部分植物中存在两种ACCase (见图1),即同质型ACCase和异质型ACCase,两种形式的ACCase都由BCCP、BC、CT组成 [1] ,但异质性CT结构域由α-CT和β-CT组成 [2] ,同质型ACCase位于同一条多肽链上,结构相对稳定;而异质性ACCase来源于不同肽链,即通过核基因对BCCP、BC、α-CT三个亚基进行编码,核基因在RNA聚合酶的作用下,通过转录合成mRNA;接下来,mRNA通过翻译生成与之配对的氨基酸序列,由此得到前体蛋白。叶绿体中的β-CT亚基是由叶绿体本身编码的,前体蛋白与β-CT亚基一起加工生产进而得到ACCase复合体,此复合体不稳定,具有高分子量,而且容易解离 [3] 。
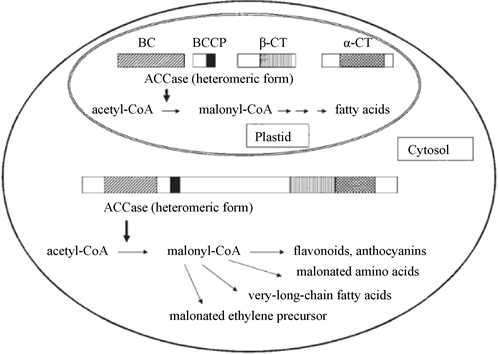
Figure 1. Two types of ACCase in plants [1]
图1. 植物中的两种ACCase [1]
2.2. ACCase的定位
异质型ACCase存在于细菌、双子叶植物及非禾本科单子叶植物,位于质体中。同质型ACCase存在于动物、酵母、藻类及大多数高等植物,位于胞质溶液中 [4] [5] 。
研究发现,在禾本科植物的质体和胞质溶液中,只有同质型ACCase [6] ,但ACCase氨基酸序列有所差别。水稻为草本稻属的一种,二倍体禾本科单子叶植物,其质体中的ACCase与胞质中的ACCase氨基酸序列同源性为73%。不可否认,质体中的ACCase对于植物脂肪酸正常的合成是不可或缺的。此外,ACCase存在方式既有同质型又有异质型的情况也是存在的,如:油菜叶绿体 [7] [8] 。
3. ACCase抑制剂类除草剂
ACCase抑制剂类除草剂出现于20世纪七十年代中期,这种除草剂常被用于双子叶植物中的杂草控制,只有少量的品种用于禾本科单子叶植物。这些除草剂可大致分为三类,即芳氧苯氧丙酸酯类(aryloxyphenoxypropionates FOPs)、环己二酮类(cyclohexanediones DIMs)和苯基吡唑啉类(phenylpyrazolin DEN),其中FOPs类和DIMs类销量最高。
3.1. 芳氧苯氧丙酸酯类(FOPs)
芳氧苯氧丙酸酯类除草剂是日本石原产业公司开发生产并进行销售的,该公司将2,4-D和二苯醚结构结合起来得到了第一个FOPs类除草剂:禾草灵(见图2),之后该类除草剂发展十分迅速、不断有新品种开发出来 [9] [10] 。代表产品有:炔草酯及氰氟草酯。
炔草酯是广谱除草剂,具有耐低温和与其它除草剂混配效果好的优势 [11] 。杂草喷药后,会产生叶片尖端变黄发黑、停止生长、皱缩死亡的表型。氰氟草酯用于水稻田,主要是通过水稻的耐受性,耐受性的基础是缺乏酯酶功能,主要是通过角质层的吸收减少和氰氟草酯代谢的增加。所有产品针对禾本科一年或多年生杂草,安全性好,选择性强。
芳氧苯氧丙酸类除草剂缺点:商品化的FOPs类除草剂通常被加工成酯类,使得其更容易被杂草所吸收;但通常酯类物质的溶解度较小,吸附性较强,因此不容易淋溶,而更容易吸收在土壤中。残留问题严重,无论对于土壤生物、水生生物还是对哺乳动物和人都会带来严重的毒害 [12] 。
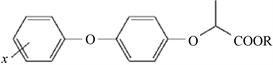
Figure 2. Molecular structure general formula of FOP herbicides [2]
图2. FOP类除草剂分子结构通式 [2]
3.2. 环己二酮类(DIMs)
环己二酮类为日本曹达公司在上世纪70年代开发,在此之前该公司将吡喃酮结构引入到杀螨剂苯螨特的类似物,研制出吡喃酮类化合物,具有除草活性。一年或多年生禾本科杂草可以被这类化合物防除,且该化合物对阔叶类植物基本无害,但芽后活性低。为此,环己二酮类化合物(见图3)被研发出来,它的芽后活性功效更为乐观。
环己二酮类除草剂具有代表性的产品是禾草灭、烯草酮。研究发现:有一种可以提高活性的方法:将含有硫原子的烷基引入到环己二酮的5号位。
环己二酮类除草剂缺点:药效受温度影响特别大。使用时应注意避免飘移到禾本科作物上 [13] 。

Figure 3. Molecular structure general formula of DIM herbicides [2]
图3. DIM类除草剂分子结构通式 [2]
4. 靶标抗性及非靶标抗性
田间杂草是导致农作物减产的最主要原因之一,杂草是具有高度竞争力的植物,它们持续地适应种植制度,造成作物的损失和破坏,而施用除草剂仍是公认的防除杂草最经济、最有效的手段。然而,不科学的施用除草剂往往会使得杂草对其产生抗性,主要是因为单一的除草剂作用的靶标唯一,且主要作用位点单一,该位点突变会使得杂草对这一类除草剂产生抗性,这对地方和全球一级的农业可持续性构成了威胁,很多杂草对ACCase抑制剂类除草剂产生抗性。
杂草对ACCase除草剂产生抗性的机制主要有两种:靶标抗性和非靶标抗性。与除草剂靶标抗性相比,由于生物化学过程固有的复杂性和杂草物种所能获得的基因组信息有限,在分子水平上阐明的非靶标抗性较少。
靶标抗性大多是单基因的,并且涉及一个点突变的目标酶。非靶标抗性可能是由植物解毒过程引起的,该过程遵循四阶段模式。第一阶段是解毒,氧化是一种典型的脱毒反应,可由P450单加氧酶或混合功能氧化酶进行。第二阶段的解毒一般是利用硫醇或糖,从而使第二阶段解毒的最终产物被第三阶段的转运者识别。第三阶段的解毒通过主动运输,将结合分子运入液泡或胞外空间。第四阶段解毒涉及进一步降解液泡或细胞外间隙的共轭分子 [14] 。
4.1. ACCase靶标基因产生抗性的氨基酸位点
ACCase抑制剂类除草剂的靶标抗性报道居多,且此类抗性能够稳定遗传给后代,通过基因的表达,使其后代能够表现出抗该除草剂的表型。非靶标抗性这一代谢途径使得小部分杂草能够存活繁衍,大部分杂草依然是通过靶标抗性进行存活繁衍。控制靶标抗性的基因片段上的保守区域发生突变的方式有:单位点突变及多位点突变,基因通过点突变,进而转录翻译得到分子结构会发生改变的蛋白质,但并不会影响其生物活性,从而使得杂草对除草剂产生抗性。
有学者发现黑麦草对ACCase抑制剂类除草剂产生抗性,此类抗性可能与1781和2096位点的氨基酸残基突变有关 [15] ,即1781位点的异亮氨酸突变成为亮氨酸;2096位点的甘氨酸突变成为丙氨酸。此外,还存在2041位点的异亮氨酸被天冬酰胺或缬氨酸所取代的情况等。ACCase抑制剂类除草剂抗性杂草氨基酸序列共有7个位置发生突变(见表1),这些氨基酸残基位置发生突变都会或多或少的使得杂草或者作物对该类除草剂产生抗性 [16] 。
4.2. ACCase抑制剂类除草剂氰氟草酯的非靶标抗性机制
有研究发现水稻对ACCase抑制剂类除草剂氰氟草酯的抗性机制为:氰氟草酯本身对于植物无毒,发挥作用是通过其酸性形式:氰氟草酸,氰氟草酯转化为氰氟草酸需通过酯酶的作用,而在水稻中此类酯酶近似于失活,故其活性形式较少。此外,已转化为活性形式的除草剂又会被谷胱甘肽转移酶处理或继续转化成无毒形式,最终对水稻无法造成伤害,起到一个保护作用 [17] [18] (见图4)。

Table 1. Amino acid substitution sites for weeds to develop resistance to ACCase herbicides
表1. 杂草对ACCase类除草剂产生抗性的氨基酸替换位点

Figure 4. Diagram of the resistance mechanism of rice to cyhalfop-butyl [19]
图4. 水稻对氰氟草酯的抗性机理图 [19]
5. 展望
在农作物的生产中,作为5大粮食作物之一的水稻,基于比较严重的杂草稻现状,急需培育出抗除草剂水稻以防控杂草稻。其中靶标抗性研究和非靶标抗性研究相比更为简单,并且不会过度涉及传统转基因问题,若应用于生产的话更容易被人接受。而且纵观除草剂市场,ACCase类抑制剂除草剂占市场份额较重,种类多且效果好,以此为靶标进行抗除草剂研究最为合适。本课题组近年也在开展ACCase靶标关键位点突变的抗性研究,期望能创制出新的抗性水稻。
基金项目
本研究由:国家重点研发计划子课题(2017YFD01005050103);天津市2018年现代农业产业技术体系创新团队建设(ITTRRS2018003)资助。
NOTES
*通讯作者。