1. 引言
我国多数地区,冬冷夏热,四季分明。夏季酷暑高温,经常威胁着植物生长发育。园林植物生产,虽然可以通过浇水灌溉来减少土壤干旱引起的水分胁迫,但是严热酷暑,却无法通过常规生产技术加以避免。特别是随着全球“温室效应”和城市“热岛效应”不断加剧,夏季高温已经成为制约园林植物生长和发育的主要环境因子 [1] 。提高植物耐热性是园林生产中亟待解决的实际问题。
5-氨基乙酰丙酸(ALA)是一种天然氨基酸,但是它不直接参与生物体内蛋白质合成,而是作为叶绿素、血红素等卟啉化合物生物合成的关键前体,参与生命代谢诸多过程。Hotta等(1997) [2] 首先提出,ALA可以促进水稻、萝卜、大麦、马铃薯、大蒜、蚕豆等多种农作物的生长,并且提高产量。Watanabe等(2000) [3] 观察到,ALA可以提高棉花的耐盐性,而且这种效应比当时所知的包括生长素、赤霉素、细胞分裂素、脱落酸、乙烯利在内的十多种植物生长调节物质效应更为显著。因而,这是一种既能提高植物抗逆性,又能增加作物产量的新型天然植物生长调节剂,在农业生产上有着广泛应用前景 [4] 。近年来,有关ALA提高植物抗逆性研究已经不再局限于盐渍,而是涉及到低温 [5] [6] 、弱光 [7] 、强光 [8] 、干旱 [9] 、水涝 [10] 、缺氮 [11] 、重金属污染 [12] 等。在植物耐热性上,首先在西瓜上观察ALA提高幼苗耐热性效应 [13] 。其后,在黄瓜 [14] 、无花果 [15] 等植物上观察类似效应。但是,ALA是否可以提高园林植物的耐热性迄今尚未见报道。本试验以金森女贞和金山绣线菊2种园林植物为材料,叶面喷施50 mg/L ALA溶液,25 d后采集离体叶片,置于
50 ℃
高温条件下,分别于0、30、60和90 min后利用M-PEA多功能植物效率仪检测叶片叶绿素快速荧光特性和820 nm光反射吸收曲线,然后利用OJIP法分析了ALA对2种园林植物叶片耐热性的影响,为其在园林生产上的应用提供理论依据。
2. 材料与方法
2.1. 材料与处理
本试验所用材料金森女贞(Ligustrum japonicum Thunb)和金山绣线菊(Spiraea japonica Gold Mound) 为多年生金叶小灌木,树龄5年,种植于泗洪县世纪公园。该地区位于北亚热带和北暖温带过渡区。冬季极端低温−23.4℃,夏季极端高温41.4℃。7月份高温天气长达7~11 d,严重影响植物生长发育。2016年4月21日用50 mg/L 5-氨基乙酰丙酸(ALA,南京禾稼春生物科技有限公司提供)溶液喷施植株叶片,直至叶面完全湿润有液滴落下为止。每种试材各设清水对照和ALA处理,每个小区面积
20 m
2
以上,重复3次,随机排列。其它正常管理。25 d后,随机采集不同小区新梢叶片,装于自封袋内,运回实验室做进一步处理分析。
将采集的叶片置于
50 ℃
恒温培养箱内,设高温处理0、30、60和90 min 4个不同处理时间,然后用M-PEA多功能植物效率仪测定叶片叶绿素快速荧光曲线和820 nm光反射荧光曲线,分析高温时间以及ALA处理对2种植物叶片光合系统I和光合系统II活性的影响,以评估其提高植物耐热性效应。
2.2. 叶绿素荧光特性测定
用英国Hansatech公司生产的多功能植物效率仪(Multifunctional Plant Efficiency Analyzer, M-PEA)测定不同处理植株叶片叶绿素快速荧光OKJIP曲线和820 nm光反射荧光吸收曲线,并按照前人方法[16]计算叶绿素荧光参数。其中,O点为初始荧光(Fo),即PSII反应中心完全开放时的荧光强度;K点为照光300 ms时荧光(Fk),J点为照光2 ms时荧光,I点为照光30 ms时荧光,Fm为最大荧光,即PSII反应中心完全关闭时的荧光强度,时间大约为在200 ms。几种主要荧光参数计算方法如下:
1) 可变荧光Fv = Fm − Fo;
2) PSII最大光化学效率φPo = (Fm − Fo)/Fm = Fv/Fm;
3) K相相对荧光Vk = (Fk − Fo)/(Fm − Fo);
4) I相相对荧光Vi = (Fi − Fo)/(Fm − Fo);
5) QA被还原的最大速率Mo = 4 × (F300 μs − Fo)/(Fm − Fo),反映PSII反应中心关闭程度;
6) 捕获的激子将电子传递到电子传递链中QA下游其他电子受体的概率Ψo = 1 − (Fj − Fo)/(Fm − Fo);
7) PSII反应中心吸收光能用于电子传递的量子产额φEo = (Fm − Fo)/Fm × Ψo;
8) 单位面积上有活性的反应中心的密度RC/CS = φPo × (Vj/Mo) × (ABS/CS);
9) 以吸收光能为基础的光合性能指数PIabs = RC/ABS × φPo/(1 − φPo) × Ψo/(1 − Ψo);
10) PSI反应中心最大光化学效率φRo = RE/ABS = TRo/ABS × (1 − Vi);
11) PSI反应中心失去电子被氧化的速率VPSI = (MRo/MR0.7 ms − MRo/MR3 ms)/(3 − 0.7);
12) PSII传递来的电子还原PSI反应中心的速率VPSII-PSI = (MRo/MR100 ms − MRo/MR7 ms)/(100 − 7)。
以上所有参数,均重复测定10张不同叶片,取平均值。
2.3. 统计分析
所得数据经SPSS20.0软件进行多因素方差分析和Duncan’s检验。当P < 0.05时,认为试验效应显著;当P < 0.01时,认为试验效应极显著。
3. 结果与分析
3.1. ALA处理对两种植物离体叶片高温后叶绿素快速荧光曲线的影响
图1展示的是ALA处理对金森女贞和金山绣线菊离体叶片经
50 ℃
高温处理不同时间后的叶绿素快速荧光曲线的影响。从中可以看出,正常条件下,未经处理的植物叶片具有典型的OKJIP变化曲线,而经ALA处理后,金森女贞叶片K、J、I和P点都有极显著上升,而金山绣线菊OJIP曲线也有一定上升,只是上升幅度不如前者那么大。因而,在非高温条件下,ALA对金森女贞叶片叶绿素快速荧光曲线的促进效应更明显。
图1还显示,高温处理对两种植物离体叶片OKJIP曲线有明显影响。在处理的30~90 min内,所有叶片O点显著上升,而且出现一个原先并不明显的K点,同时J、I和P点都显著下降,甚至消失,说明高温处理显著抑制PSII反应中心供体侧放氧复合体(Oxygen evolving complex, OEC)活性,并因此抑制受体侧光合电子传递。
未经ALA处理的对照叶片,经高温处理后JIP几乎一蹶不起;而ALA处理叶片经
50 ℃
高温胁迫30 min的叶绿素荧光在100 ms后仍然有一定的上升。所有经ALA处理叶片荧光曲线均明显高于对照,暗示着该药剂处理可以提高热胁迫下两种园林植物叶片PSII反应中心荧光活性。

Figure 1. Effects of ALA treatment on chlorophyll fast fluorescence curves of detached leaves of L. japonicum and S. japonica after 50˚C high temperature stress for different times
图1. ALA处理对金森女贞和金山绣线菊离体叶片经
50 ℃
高温处理不同时间后的叶绿素快速荧光曲线的影响
3.2. ALA处理对两种植物离体叶片高温后PSI反应中心820 nm光反射荧光曲线的影响
图2显示,金森女贞和金山绣线菊叶片820 nm光反射荧光曲线呈V字型,即从0.7 ms (JIP时间)到大约10 ms间,MRo/MR荧光迅速下降,代表着照光初期PSI反应中心P700失去电子被氧化成P700*过程;而从那以后,MRo/MR荧光缓慢上升,代表着P700通过自身环式电子传递链或来自PSII反应中心非环式电子传递链得到电子被还原的过程。实际上,在MRo/MR下降阶段,一方面存在着P700被氧化,另一方面,也存在着得到电子被还原,但是由于氧化速率大于还原速率,因而,氧化过程占优势。当MRo/MR达到最低值时,失去电子与获得电子达到平衡状态,但从那以后,来自于PSII反应中心的电子增多,P700逐渐呈还原状态,MRo/MR荧光曲线上升。当光照时间达100 ms以上时,PSII反应中心完全被氧化,处于完全关闭状态,它能够传递给PSI的电子逐渐下降,MRo/MR曲线又开始下降。因而,10 ms附近的MRo/MR最低值代表着PSI反应中心能够被氧化的电子载体量,而100 ms以后的MRo/MR最高值代表着PSII反应中心能够传递给PSI末端电子受体的电子负荷量。
从图2可以看出,ALA处理后金森女贞和金山绣线菊叶片MRo/MR最低值均显著低于对照,而最高值都高于或略高于对照,说明ALA处理不仅可以提高两种园林植物叶片PSI电子供体量,也能提高PSII反应中心提供给PSI反应中心的电子负荷量。其中,ALA对金森女贞的促进作用大于金山绣线菊。
高温胁迫极显著提高了MRo/MR的最低值,同时也降低了MRo/MR最高值,说明两个光合系统活性都受到伤害。与金山绣线菊相比,金森女贞PSI活性对高温胁迫更为敏感,因为
50 ℃
高温处理30 min时,金山绣线菊叶片MRo/MR曲线下降后仍然有比较明显的上升,而金森女贞的上升过程相对比较弱。类似地,ALA处理后金山绣线菊叶片MRo/MR曲线也有明显的下降-上升过程,而金森女贞的反应较弱。这些数据反映出,ALA处理显著提高未经高温胁迫的金森女贞叶片PSI活性,但是ALA处理的金山绣线菊叶片具有更强的耐热性。
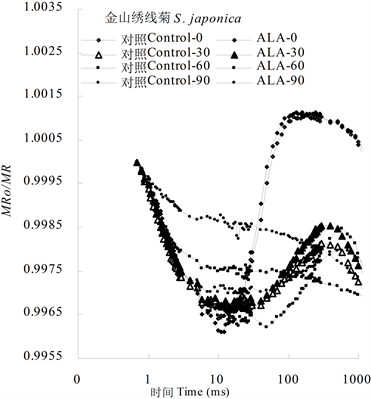
Figure 2. Effect of ALA treatment on the modulated 820 nm reflection curves of leaves of L. japonicum and S. japonica after 50˚C high temperature stress for different times
图2. ALA处理对金森女贞和金山绣线菊离体叶片经
50 ℃
高温处理不同时间后的820 nm光反射动力学曲线的影响
3.3. ALA处理对两种植物离体叶片叶绿素荧光参数的影响
3.3.1. 初始荧光Fo
图3显示,高温胁迫后,两种植物初始荧光Fo显著上升,而ALA处理进一步导致Fo上升。对于金森女贞来说,当高温超过60 min,ALA处理叶片Fo极显著高于对照。对于金山绣线菊来说,Fo也表现出类似趋势,但只有高温90 min时,ALA处理叶片Fo才极显著高于对照,而在此之前,两者间没有显著差异。高温90 min时,ALA处理Fo明显高于对照的原因,一方面是对照叶片Fo下降,另一方面是ALA处理叶片Fo继续上升。这说明,Fo上升是植物叶片光合膜对高温胁迫的反应。也许,只有有活性的光合膜才能对高温胁迫产生这种生物学反应。未经ALA处理的金森女贞叶片在高温处理60 min后已经失去对高温胁迫的响应,其Fo与高温30 min的没有显著差异,而ALA处理叶片Fo仍然大量上升,暗示着这些叶片的光合膜仍然具有高温热反应。类似情况存在于金山绣线菊叶片。高温处理90 min时,对照叶片Fo已经失去对高温的反应能力,而ALA处理叶片的Fo继续上升,说明其仍然保持着生物学活性。
3.3.2. 最大荧光Fm
图4显示,高温处理极显著降低金森女贞和金山绣线菊叶片最大荧光值Fm,其中
50 ℃
高温处理30 min对金森女贞的效应与60或90 min的相似,Fm值下降一半以上,但金山绣线菊Fm下降程度与高温处理时间有关。时间越长,下降越多,两者呈直线关系(r = 0.958, P < 0.05)。这说明该物种对高温胁迫具有较强耐受力。高温处理90 min,其叶片Fm仍然对高温胁迫有响应。但对金森女贞来说,高温处理30 min后叶片Fm已经丧失了对热胁迫响应。再次说明,金森女贞对高温胁迫更敏感。
从ALA处理效应上看,高温处理前,ALA处理的金森女贞叶片Fm比对照高出60% (P < 0.01),而金山绣线菊叶片仅高出7%,未达到差异显著水平(P > 0.05)。随着高温时间延长,ALA对金森女贞叶片Fm的促进效应均维持在45%左右,没有明显变化;但ALA对金山绣线菊Fm的促进效应随着高温处理时
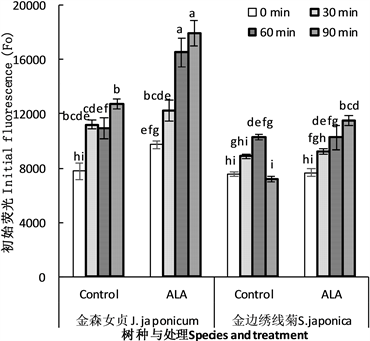
Figure 3. Effect of ALA treatment on the initial fluorescence (Fo) of detached leaves of J. japonicum and S. japonica after heat stress. The same lowercases above the histogram represent no significant difference at P = 0.05 level. The same as below
图3. ALA处理对金森女贞和金山绣线菊离体叶片高温胁迫后叶绿素快速荧光的初始荧光(Fo)的影响。柱形图上相同小写字母表示在P = 0.05水平上差异不显著,下同

Figure 4. Effect of ALA treatment on the maximum fluorescence (Fm) of leaves of J. japonicum and S. japonica after heat stress
图4. ALA处理对金森女贞和金山绣线菊离体叶片高温后叶绿素快速荧光的最大荧光(Fm)的影响
间延长而呈直线上升(r = 0.966, P < 0.05)。这说明,对于两种耐热性不同的园林植物来说,ALA促进金山绣线菊耐热性效应高于金森女贞,ALA对前者耐热性调节的幅度也大。
3.3.3. 可变荧光Fv
与Fm相似,ALA处理诱导两种植物叶片叶绿素可变荧光Fv显著上升(F > 0.01),而高温处理导致Fv迅速下降。对金森女贞来说,未经高温处理的叶片所体现出来的ALA处理后增幅达70%以上。然而,与
50 ℃
高温30 min相比,这个增加幅度并不大,因为在高温30 min后,金森女贞叶片Fv增幅达118%,表明在这种胁迫条件下,ALA可以表现出良好的调节活性。但是,随着高温时间进一步延长,ALA诱导的Fv增幅又下降为46%~62%。与此不完全相同的是,ALA诱导金山绣线菊叶片Fv增幅最大的是高温处理60 min,增幅为198%。另外,高温处理90 min的叶片Fv增幅也达117% (图5)。显然,ALA对常温条件下金山绣线菊叶片Fv的影响不大,但在高温胁迫下,尤其是相对较长时间的高温胁迫下,可以体现出更好的调节效应。
3.3.4. PSII反应最大光化学效率(Fv/Fm)
Fv/Fm,或者φPo是暗适应叶片最大光化学效率。高温处理前,ALA对两种园林植物叶片φPo没有显著影响。高温胁迫30 min后,两种园林植物叶片φPo都显著下降,但ALA处理叶片能保持较高水平,其中金森女贞显著高于对照(P < 0.05)。高温胁迫60或90 min,经ALA处理的金山绣线菊叶片φPo显著高于对照,但金森女贞叶片φPo与对照没有差异。这说明,经ALA处理的金森女贞叶片可以忍受短时间的高温胁迫,而金山绣线菊则可以忍受更长时间的高温逆境。
3.3.5. K相相对荧光(Vk)
K相对荧光Vk是指暗适应叶片在强光照射300 ms时的相对荧光值,所反映的是PSII反应中心供体侧放氧复合体(OEC)相对受抑程度。与Wk相似,Vk越高,说明OEC受抑程度越大。从图6可以看出,随着高温时间延长,两种植物叶片Vk显著上升,特别是金森女贞,高温处理30 min后叶片Vk极显著高于处理前,而且与其后高温60或90 min没有显著差异(图7),说明
50 ℃
高温处理30 min就已经伤害到PSII反应中心供体侧OEC活性,因而,即使其后延长高温时间,也已经不再产生新的伤害。但是金山绣线菊叶片Vk随着高温时间延长而逐渐上升,表明它对高温逆境的响应是一个逐渐发展的过程。这也说明了金山绣线菊对高温的忍受力高于金森女贞。
从另一方面看,ALA处理显著降低两种植物叶片Vk (F > 0.01),但是两种植物间的响应模式有所不同。对金森女贞来说,未经高温或高温处理30 min的叶片对ALA处理效应显著,而更长时间高温处理则丧失处理效应。对金山绣线菊来说,未经高温处理或者高温处理30 min叶片的ALA处理效应较小,而高温处理60或90 min的处理效应越发明显。也就是说,随着高温处理时间的延长,ALA处理效应越
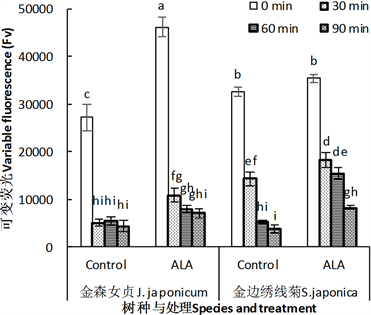
Figure 5. Effect of ALA treatment on the variable fluorescence (Fv) of leaves of J. japonicum and S. japonica after heat stress
图5. ALA处理对金森女贞和金山绣线菊离体叶片热胁迫后叶绿素快速荧光的可变荧光(Fv)的影响

Figure 6. Effect of ALA treatment on the relative fluorescence of K phase (Vk) of detached leaves of J. japonicum and S. japonica after heat stress
图6. ALA处理对金森女贞和金山绣线菊离体叶片高温胁迫后K相相对荧光(Vk)的影响
高。因而,ALA处理可以在短期高温条件下保护金森女贞叶片PSII反应中心供体侧OEC活性,也可以在较长时间高温逆境中保护金山绣线菊叶片PSII反应中心OEC活性。
3.3.6. PSII反应中心关闭最大速率(Mo)
与Vk相似,高温逆境导致两种园林植物叶片PSII反应中心受体侧QA被完全还原的速率(Mo)迅速上升(图8),意味着PSII反应中心完全关闭的速度加快。但是,金森女贞与金山绣线菊的响应不完全相同。
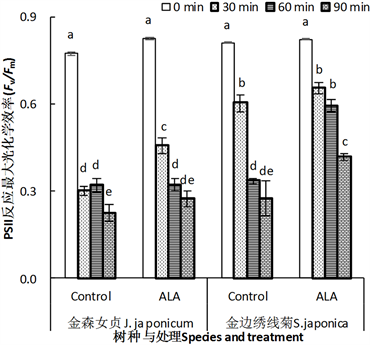
Figure 7. Effect of ALA treatment on the maximum photochemical efficiency (Fv/Fm = ΦPo) of PSII reaction center of leaves of J. japonicum and S. japonica after heat stress
图7. ALA处理对金森女贞和金山绣线菊离体叶片高温胁迫后PSII反应中心最大光化学效率(Fv/Fm = ΦPo)的影响
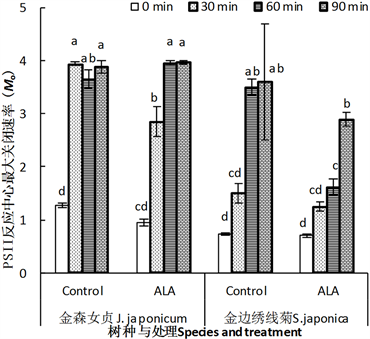
Figure 8. Effect of ALA treatment on the maximum close rate of PSII reaction center (Mo) of detached leaves of J. japonicum and S. japonica after heat stress
图8. ALA处理对金森女贞和金山绣线菊离体叶片经高温处理后PSII反应中心最大关闭速率(Mo)的影响
对前者来说,30 min高温处理的Mo与60或90 min高温处理的Mo几乎完全相等,说明30 min高温导致了叶片QA被完全还原。对金山绣线菊来说,高温处理60 min才会导致Mo显著高于短时间高温处理或者未经高温处理。这也说明金山绣线菊叶片QA被还原对高温的忍耐力强于金森女贞。
ALA处理抑制高温诱导的叶片Mo上升效应在两种园林植物中表现模式存在差异。对金森女贞来说,ALA抑制未经高温或者短时间高温处理的叶片Mo上升,但是对高温60 min及其以上时间的叶片没有效应。对金山绣线菊来说,未经高温处理或者高温处理90 min叶片中观察不到ALA处理效应,但是,在高温处理30或60 min时,ALA处理效应显然存在,尤其是在高温60 min的叶片。
3.3.7. PSII捕获的光能用于光合电子传递的量子产额(φEo)
图9显示,高温处理导致两种园林植物叶片PSII反应中心捕获的光能用于光合电子传递的量子产额φEo下降,但是,两者之间存在着明显差异。对金森女贞来说,
50 ℃
高温处理30 min,φEo下降68%,即使延长高温时间,也是保持相似的下降幅度。但对金山绣线菊来说,高温处理30 min,φEo才下降15%;若高温处理60 min,φEo下降83%。再次说明,金山绣线菊叶片φEo对高温的忍受能力高于金森女贞。
ALA处理可以显著提高两种园林植物叶片φEo对高温逆境的忍受力。其中,金森女贞叶片在未经或30 min高温时,ALA处理叶片φEo显著高于对照,而金山绣线菊叶片经高温处理后φEo均显著高于对照(图9)。
3.3.8. PSII有活性反应中心密度(RC/CS)
图10展示的是两种园林植物叶片PSII有活性反应中心的密度(RC/CS)。从中可以看出,
50 ℃
高温处理30 min后,金森女贞叶片RC/CS下降了79%,金山绣线菊下降了62%。随着高温时间延长,金森女贞RC/CS几乎没有继续下降,而金山绣线菊下降到原初值到80%左右便不再下降。说明高温胁迫导致两种植物叶片RC/CS下降程度相近,只是金森女贞在短时间高温胁迫后就降低到最低程度,而金山绣线菊需要更长时间才降低到最低程度。这说明,后者更耐热。
ALA处理显著提高两种植物叶片PSII有活性反应中心对高温的适应性,其中,金森女贞对ALA处

Figure 9. Effect of ALA treatment on the quantum yield for electron transport (φEo) of detached leaves of J. japonicum and S. japonica after heat stress
图9. ALA处理对金森女贞和金山绣线菊离体叶片经高温处理后PSII反应中心用于电子传递的量子产额(φEo)的影响
理效应比较稳定。无论是经或未经高温处理的叶片,RC/CS比对照高出40%~87%。但对金山绣线菊来说,未经或仅经高温处理30 min的叶片,RC/CS仅增加5%~10%,没有达到差异显著水平(P > 0.05),只有在高温处理60 min时,ALA处理增幅才达到78%,显著高于对照(P < 0.05)。
3.3.9. PSI反应中心最大光化学效率(φRo)
图11显示,两种园林植物叶片光合系统I反应最大光化学效率对高温处理存在着明显差异,其中金
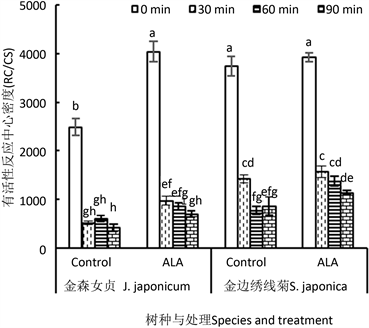
Figure 10. Effect of ALA on the active PSII reaction center density (RC/CS) of detached leaves of J. japonicum and S. japonica after heat stress
图10. ALA处理对金森女贞和金山绣线菊离体叶片高温处理后有活性反应中心密度(RC/CS)的影响
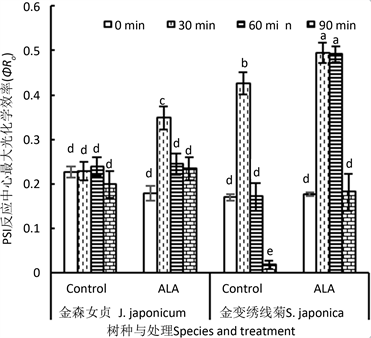
Figure 11. Effect of ALA on the maximum photochemical efficiency to reduce the end electron receptors of PSI reaction centers (φRo) of detached leaves of J. japonicum and S. japonica after heat stress
图11. ALA处理对金森女贞和金山绣线菊离体叶片经高温处理后PSI反应中心最大光化学效率(φRo)的影响
森女贞叶片φRo对
50 ℃
高温处理没有明显反应,而金山绣线菊叶片φRo在高温处理30 min时显著上升,但在高温90 min后则极显著低于未处理对照,表明该物种叶片PSI反应中心对高温胁迫有相当灵活的调节能力。短时间高温胁迫诱导PSI活性上升,长时间高温逆境导致φRo下降。φRo下降是高温胁迫引起的伤害结果。
ALA处理对两种植物φRo的影响也存在显著差异。未经高温处理的金森女贞叶片φRo对ALA处理没有反应,高温30 min处理诱导φRo上升;更长时间高温后,ALA处理效应消失。对金山绣线菊来说,未经高温处理叶片φRo没有因ALA处理而出现明显效应,而高温处理不同时间后,所有叶片φRo均显著上升。其中,高温处理30 min叶片φRo增加16%,高温处理60或90 min则分别比对照高出1.87倍和8.69倍。
3.3.10. 以吸收为基础的光合性能指数(PIabs)
图12显示,两种园林植物叶片以吸收为基础的光合性能指数PIabs对高温胁迫非常敏感,只要
50 ℃
高温处理30 min,金森女贞叶片PIabs下降99%,金山绣线菊叶片也下降78%。如果高温延长到60或90 min,金山绣线菊PIabs下降99.5%。这些现象一方面说明,PIabs的确是一个非常敏感的高温伤害指标,同时也证明,金山绣线菊叶片对高温的忍受能力强于金森女贞。
ALA处理显著提高两种植物PIabs,但两种植物之间存在着差异。对金森女贞来说,ALA处理提高了未经或仅经高温30 min叶片的PIabs,增幅达1.35和6.80倍。但从统计角度看,只有前者差异达到P = 0.05水平。对金山绣线菊来说,ALA提高所有叶片PIabs,其中未经高温处理叶片增加16%,经高温处理60 min叶片的PIabs增加35.78倍。两者与对照相比,差异达到P = 0.01显著水平。
3.3.11. PSI反应中心氧化还原速率(VPSI和VPSII-PSI)
图2已经列出来的“V”字型820 nm调制反射曲线,其MRo/MR下降阶段为PSI反应中心失去电子被氧化过程,上升阶段为PSII反应中心传递来的电子还原PSI反应中心过程。以0.7 ms到3 ms间MRo/MR下降值,可以计算出PSI反应中心被氧化的速率VPSI (见图13A);以7 ms到100 ms间MRo/MR上升值,

Figure 12. Effect of ALA on the capacity index based on absorption (PIabs) of detached leaves of of J. japonicum and S. japonica after heat stress
图12. ALA处理对金森女贞和金山绣线菊离体叶片经高温处理后以吸收为基础的性能指数(PIabs)的影响

Figure 13. Effect of ALA on the oxidation (VPSI) and reduction rates (VPSII-PSI) of PSI reaction center of detached leaves of J. japonicum and S. japonica after heat stress
图13. ALA处理对金森女贞和金山绣线菊离体叶片经高温处理后PSI反应中心氧化速率(VPSI)和还原速率(VPSII-PSI)的影响
可以计算出PSII传递过来电子还原PSI反应中心的速率VPSII-PSI (见图13B)。
从图13A上看,高温胁迫导致两种植物叶片VPSI下降,而ALA处理可以提高植物PSI反应中心的耐热性。就常温对照而言,经ALA处理的金森女贞叶片VPSI虽然高于对照,但从统计学上看,两者差异不显著。当叶片经50℃高温处理30或60 min时,ALA处理叶片VPSI上升,对照下降,两者呈现出极显著差异(P < 0.01)。对金边绣线菊来说,常温或短时间高温胁迫时,ALA对VPSI的影响不大,而在60 min,特别是90 min高温胁迫时,ALA处理叶片VPSI显著高于对照。从而说明,ALA对两种园林植物叶片PSI反应中心耐高温能力有显著促进效应。
从图13B可以看出,从PSII反应中心传递来的电子还原PSI反应中心的速率(VPSII-PSI)比PSI自身氧化速率(VPSI)低1个数量位,但是,VPSII-PSI对高温的敏感性却远远高于VPSI。无论是金森女贞还是金边绣线菊,50℃高温处理后,VPSII-PSI迅速下降,而且金森女贞下降速度大于金边绣线菊,未经ALA处理叶片大于经处理叶片。这说明两个光合系统间电子传递对高温胁迫非常敏感,而ALA处理可以提高两种园林植物VPSII-PSI对高温胁迫的敏感性。
4. 讨论
4.1. 植物耐热性与叶绿素荧光特性
金森女贞和金山绣线菊均是园林生产中常用的金黄叶彩色树种。其中前者原产于日本,在我国种植有十多年历史;后者来自于北美,喜欢光照温暖湿润气候和肥沃土壤,而且耐寒性较强 [17] [18] ,适宜在长江以北多数地区栽培。有人提出,金森女贞比金叶女贞更耐高温 [19] ,适应我国南方夏季高温环境。但也有人 [20] 提出,金森女贞属于最不耐热的一类园林植物,而金山绣线菊的耐热性尚未见报道。本试验利用离体叶片经高温胁迫后再用M-PEA检测叶绿素荧光特性,所获证据表明,金森女贞的耐热性的确不如金山绣线菊。一般植物叶片经充分暗适应后照光,其叶绿素可以发出一系列荧光,形成典型的OJIP荧光变化。但是,金森女贞的OJIP变化幅度明显不如金山绣线菊(图1)。特别是在
50 ℃
高温处理后,金森女贞荧光曲线除了K点上升外,其它的JIP点几乎不再起来,且恢复得比金山绣线菊少。由此看来,金森女贞叶片(离体)对高温的忍受力不如金山绣线菊。从PSI反应中心820 nm光反射吸收荧光曲线上看,高温处理后,金森女贞的MRo/MR最低值迅速上升,而最高值迅速下降,说明其PSI反应中心活性很容易失活。从其它叶绿素荧光参数(见下文)上看,两种植物的特性差异也明显存在。
不同的荧光参数代表着不同的生物学意义。比如,Fo,代表着光合膜结构的稳定性。一般认为Fo越低越好。逆境胁迫导致Fo上升,是一种伤害症状 [21] [22] 。但是,从本试验结果看,
50 ℃
高温胁迫30 min导致金森女贞叶片Fo显著上升;高温时间再延长,Fo却不再有明显反应。相反,金山绣线菊叶片Fo在高温处理60 min时仍然处于上升状态。这一结果说明,不能简单地以Fo高低来判定高温对光合膜的影响。Fo变化是一种正常的生理反应。但若不再变化,那么,光合膜可能受到了不可逆的伤害。小麦上,如果将叶片以
2 ℃
/min速率缓慢加热上升,Fo会在某一温度临界点突然上升 [23] 。为此,作者提出,Fo的这一温度变化临界点可以作为判定该物种耐热性的指标。本试验没有研究梯度升温效应,但是从高温处理的时间效应上看,两种园林植物叶片Fo都存在着先上升,然后不变或者下降的变化动态(图3)。我们认为,可以把Fo这个上升点作为物种耐热性的关键点,也可以将Fo不变或下降的时间点作为该物种耐热性判定的依据。也许后者更为可靠。因而,我们应该动态而不是静态地看待Fo这一荧光参数。
除了Fo之外,Fm、Fv、F300 ms(或者叫Fk)、Fj、Fi等都是非常重要的荧光数据点,经常用来探讨植物叶片PSII反应中心供体侧和受体侧活性。然而,在本试验研究中,常用的生理指标Wk (K相占J相的相对荧光)、Vj、Vi以及Ψo等都没有表现出前人报道的那种特有规律,只有Vk (K相相对荧光)和Fv/Fm等表现出对热胁迫的良好响应。因而,图4~图8 (分别为Fm、Fv、Fv/Fm、Vk和Mo)均反映出两种植物对热胁迫的响应特征。Fm是最大荧光,Fv是可变荧光,Vk代表PSII反应中心供体侧放氧复合体活性受抑制程度,Fv/Fm代表PSII反应中心最大光化学效率,Mo反映PSII反应中心受体侧QA被完全还原的速率。因而,Fm、Fv、Fv/Fm越高,说明光化学能力越强,而Vk和Mo越小,说明光合原初反应活性越高。从本试验研究结果看,这些荧光参数对高温胁迫的反应与Fo相似,金森女贞离体叶片由于耐热性弱于金山绣线菊,因而,在
50 ℃
高温处理30 min后,便迅速下降,而金山绣线菊耐热性较强,在高温处理的30~90 min内,随着处理时间延长,而呈现有规律性的变化。这些结果说明,这些荧光参数能够较好地反映出物种叶片在高温胁迫下的伤害程度。
除此之外,PSII反应中心捕获的光能用于电子传递的量子产额φEo (图9)、有活性的反应中心密度RC/CS (图10)、PSI反应中心用于还原受体侧末端电子受体的量子产额φRo (图11)、以吸收为基础的光合性能指数PIabs (图12)以及PSI反应中心氧化还原速率VPSI和VPSII-PSI (图13)等,都能很好地反映出金森女贞和金山绣线菊叶片对高温胁迫的响应,可以作为植物耐热性评判的重要指标。
4.2. ALA与园林植物耐热性
5-氨基乙酰丙酸(ALA)是一种天然氨基酸,同时具有多种生长发育调节活性 [24] 。有关ALA提高植物抗逆性的研究已经很多报道 [2] [5] [6] [7] [9] [10] [16] [25] [26] [27] 。它们涉及到植物耐盐性、耐寒性、耐弱光性、耐旱性、耐涝性等,但是与植物耐高温性的研究报道相对较少 [14] [15] 。ALA与园林植物耐热性的关系迄今未见有报道。本试验以金森女贞和金山绣线菊为材料,研究了叶面喷施ALA溶液25 d后离体叶片的耐热性,发现ALA能显著提高植物耐热性。这不仅表现在它对PSII反应中心供体侧放氧复合体活性的维护效应(Vk极显著下降,图7),同时也表现在对PSII反应中心最大光化学效率(Fv/Fm,图6)以及受体侧QA开放程度(Mo较低,图8)。因而,ALA对高温胁迫下园林植物叶片PSII反应中心的保护效应是显著的。而且,这种保护作用与其它逆境如低温 [16] 、弱光 [8] 、水涝 [10] 等一样,都与PSII反应中心有关。本试验还观察到,ALA处理提高金森女贞和金山绣线菊叶片耐热性还与其提高叶片PSI反应中心还原末端电子受体活性φRo有关(图11)。这与谢荔等 [28] 在葡萄上观察的结果相吻合。他们提出,ALA处理可以诱导植物叶片抗氧化酶活性提高,其中SOD等位于叶绿体光合系统I附近,可以清除光合电子传递链中产生的过剩的活性氧,通过水-水循环 [29] ,消除活性氧对光合系统的伤害效应,保持电子传递链流畅,提高植物光合效率。因而,在园林植物中推广应用ALA,可以提高植物耐高温胁迫能力,提高植物光合性能,促进植株生长,在炎热的夏季达到美化和绿化效果。
参考文献