摘要: WUSCHEL和CLAVATA3是拟南芥茎尖分生组织调控网络中的两个关键基因,它们在茎尖分生组织中的相邻区域表达,在转录水平上存在相互调控关系。现有观点认为WUS蛋白对CLV3的表达主要具激活的功能,Perales等人的近期研究结果发现WUS蛋白可根据其浓度对CLV3转录有激活或抑制的作用。该发现不仅提供了一个WUS与CLV3相互调控的新模式,同时也可解释为什么CLV3在WUS的表达区内没有表达。
Abstract:
WUSCHEL and CLAVATA3 are two key genes in the regulatory network of shoot apical meristems in Arabidopsis. They are expressed in the adjacent regions of the shoot apical meristem and regulate each other at the transcriptional level. It is believed that WUS protein can activate the expression of CLV3. Recent studies by Perales et al. found that WUS protein could activate or inhibit the transcription of CLV3 according to its concentration. This finding not only provides a new mode of interaction between WUS and CLV3, but can also explain why CLV3 is not expressed in the WUS-expressing zone.
高等植物中的茎尖分生组织是地上器官形成与生长发育的主要场所和控制中心 [1] [2],为一个半球状的穹型结构,内含多层细胞。位于最顶端的2层细胞通常进行垂周分裂,称为原套(tunica);原体(corpus)位于原套之下,由多层细胞所构成,其中的细胞可沿多个平面进行分裂。在原体的中央有一团干细胞(stem cell),称为组织中心(organising center)。组织中心内的干细胞分裂活动不旺盛,但对茎尖分生组织的结构与功能的维持有至关重要的作用。
WUSCHEL (WUS)和CLAVATA3 (CLV3)是拟南芥茎尖分生组织调控网络中的关键基因 [2] [3] [4]。WUS编码一个同源异型结构域蛋白(homeodomain protein)家族成员,在组织中心的细胞中特异性表达 [2] [3] [5] [6]。CLV3编码一个小分子分泌型蛋白,其表达区是位于组织中心上方的3层细胞 [4] [7] [8]。CLV3和WUS功能缺失的突变体在茎尖呈现相反的表型。wus突变体缺失茎尖分生组织或茎尖分生组织功能异常,茎尖生长缓慢,缺乏主茎而有多个分枝,每个分枝点形成莲座叶等;clv3突变体则呈现出茎尖和茎尖分生组织膨大,多心皮等表型 [2] [4] [9]。
WUS和CLV3在转录水平上相互调控(图1) [10]。WUS蛋白通过细胞间移动进入CLV3的表达区,然后与CLV3启动子中的特定位点结合对CLV3的转录产生正调控作用;而CLV3蛋白通过与其受体CLV1/CLV2的共同作用反过来对WUS的转录产生抑制作用,由此限制后者的表达区向上扩展。WUS和CLV3之间的反馈调控关系能较完满地解释WUS不在CLV3的表达区内表达,以及wus和clv3突变体在茎尖所呈现的相反表型的原因,因而被广泛接受。但是该调控机理无法解释为什么在WUS的表达区内CLV3不表达的现象。
Perales等人 [10] 的近期研究结果为上述现象提供了一个比较合理的解释。他们发现WUS蛋白可依据其浓度与CLV3基因启动子中的特异性结合位点以单体或双聚/多聚体的方式结合,在低浓度的细胞中主要以单体的形式结合,这一结合形式导致对CLV3的转录的正调控(激活)作用;在高浓度的细胞中WUS蛋白则主要以双聚体/多聚体的形式结合,该结合形式导致对CLV3的转录的负调控(抑制)作用(图2)。因此,WUS蛋白可根据其浓度以不同形式与CLV3启动子中特异性位点结合,并由此行驶激活或抑制CLV3转录的功能。这一新发现的调控机理不仅为WUS和CLV3之间的调控关系提供了新的调控模式,同时也可解释CLV3为什么不在WUS的表达区表达。
 注:WUS的表达区用淡绿色标注,CLV3的表达区用橘黄色标注。WUS和CLV3间的调控关系用不同颜色和类型的线条标示,红色线条表示负调控关系,绿色箭头示正调控关系。右侧方框中展示在CLV3表达区中WUS对CLV3转录的正调控作用的实现途径
注:WUS的表达区用淡绿色标注,CLV3的表达区用橘黄色标注。WUS和CLV3间的调控关系用不同颜色和类型的线条标示,红色线条表示负调控关系,绿色箭头示正调控关系。右侧方框中展示在CLV3表达区中WUS对CLV3转录的正调控作用的实现途径
Figure 1. The expression domains of and transcriptional inter-regulations between WUSCHEL (WUS) and CLAVATA3 (CLV3) in the shoot apical meristems of Arabidopsis
图1. WUSCHEL与CLV3在拟南芥茎尖分生组织中的表达区域和相互调控
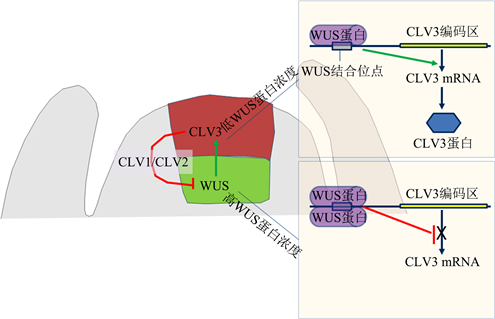 注:WUS蛋白在CLV3表达区和WUS表达区对CLV3的相反的转录调控。在CLV3表达区,WUS蛋白通过细胞间移动而来,浓度相对较低,主要以单体与CLV3启动子中的特异性结合位点结合,对CLV3转录主要呈现为正调控;在WUS表达区,WUS蛋白浓度相对较高,主要以双聚体或多聚体的形式与CLV3启动子中的特异性结合位点结合,对CLV3转录主要呈现为负调控
注:WUS蛋白在CLV3表达区和WUS表达区对CLV3的相反的转录调控。在CLV3表达区,WUS蛋白通过细胞间移动而来,浓度相对较低,主要以单体与CLV3启动子中的特异性结合位点结合,对CLV3转录主要呈现为正调控;在WUS表达区,WUS蛋白浓度相对较高,主要以双聚体或多聚体的形式与CLV3启动子中的特异性结合位点结合,对CLV3转录主要呈现为负调控
Figure 2. A new model for the dosage-dependant regulation of CLV3 transcription in by WUS in Arabidopsis shoot apical meristem
图2. 拟南芥茎尖分生组织中WUS蛋白对CLV3基因转录的剂量调控新模式
基因调控中的剂量效应在所有多细胞生物中普遍存在,但同一调控因子因其自身的剂量变化导致相反调控结果的例子并不多。Perales等人发现的WUS蛋白对CLV3的剂量调控机制是否存在于WUS蛋白对其它目标基因的调控中,或其它调控蛋白对其目标基因的调控关系中还有待进一步的研究。
基金项目
本研究得到了重庆师范大学人才基金(12XLR36)的资助。