1. 引言
光合作用是生态系统碳氧循环与植物制造养分的重要途径,而由于林木冠层对光的截获能力不同,导致光合特性呈现一定的空间分布 [1] 。国外学者对林木冠层光合作用的研究较早,Sakowska等(2018)和O’Rourke等(1981)研究发现,冠层光合作用呈现出一定的空间异质性 [2] [3] 。Suwa等(2007)研究发现,全年冠层上部叶片光合速率为冠层下部的3~16倍 [4] ,冠层光合作用与光照、叶龄等因素有关 [5] 。由于冠层内不同位置叶片的光合能力不同且叶量的垂直分布并非均匀分布,因此冠层内不同位置枝叶的光合产量及其对树干生长的贡献也存在差异。研究光合特性在冠层的变化规律,对于在冠层水平上揭示植物固碳能力和估算植物生产力具有重要意义 [6] [7] 。
光合速率不仅是植物生长发育的基础,也是植物有机物质积累的过程,是反映植物生长变化的一项重要的生理指标,它受叶片内部气孔活动状态、蒸腾速率、外界空气温度和空气相对湿度等因子的影响 [8] ,如针对单株树木,其冠层内不同位置叶片的光合特性往往存在一定差异 [9] [10] ,引起该差异的主要因素包括:光照强度、温度和湿度等环境因子 [11] 以及叶片含氮量和比叶重等特征因子 [12] 。通过测定植物光合特性,研究净光合速率与生理生态因子间的关系,为植物生长调控提供理论依据。
胡杨(Populus euphratica Oliv.)为高大落叶乔木,雌雄异株,具有耐高温、耐盐碱、耐湿、抗干旱、抗风沙的特性,是新疆干旱荒漠区的主要建群种和抗逆种质资源。胡杨林不仅在荒漠景观中占有特殊地位,也起着维护生态安全的屏障作用,在塔里木河下游沙漠化防治和绿洲生态安全保护中有不可代替的作用。胡杨生理生态适应性研究主要集中在不同生态条件下光合生理特性的比较。研究表明,胡杨光合速率日的变化规律或为“单峰”曲线 [13] [14] [15] [16] 或为“双峰”曲线 [17] [18] ,各生理、生态因素对胡杨光合过程的影响是相互且复杂的 [14] [17] [19] 。但目前,关于胡杨不同冠层光合作用的研究较少 [20] ,特别是关于胡杨雌雄株不同冠层光合生理差异的研究尚未见报道,而胡杨雌雄株在生长、物候等方面均表现出明显的差异 [21] 。本研究以胡杨雌雄为研究对象,旨在揭示雌雄株光合生理特性在冠层上变化的差异及其内外影响因素,为进一步揭示胡杨雌雄株的生态适应性机制以及林木定向培育奠定理论基础。
2. 研究地点与研究方法
2.1. 研究区概况
本实验样地位于塔里木盆地西北缘(81˚17'56.52E, 40˚32'36.90N)新疆阿拉尔市人工胡杨、灰胡杨混交林,受塔克拉玛干沙漠的影响,属暖温带极端大陆性干旱荒漠气候,地表蒸发强烈,全年降雨稀少,年均蒸发量1876.6~2558.9 mm,年均降水量40.1~82.5 mm,海拔997~1340 m,年平均气温10.7℃,≥5℃积温4325℃,全年太阳总辐射量141 Kcal/cm2,无霜期平均210 d,年均风速2.1 m/s。
2.2. 研究方法
于2021年10月2日,在研究区选择长势良好且健康的20径阶胡杨雌雄株各3棵为样株,胡杨雌雄样株基本信息见表1。在样株树高2 m、4 m、6 m、8 m、10 m处,选取东南西北四个方向各三个当年生枝,从枝条基部开始的第3、4节位叶进行光合参数的测定,每片叶测定重复3次。同时用ImageJ软件进行测试叶片叶面积的测量。

Table 1. Basic information of female and male samples of Populus euphratica
表1. 胡杨雌雄样株的基本信息
注:同一列不同字母表示显著性差异(P < 0.05)。
使用Li-6400便携式光合仪透明叶室分不同冠层测定胡杨雌雄株的光合日动态及环境因子。实验测定步长2 h,时间为9:30~19:30。测定参数主要包括:净光合速率(Pn/μmol CO2∙m−2∙s−1)、蒸腾速率(Tr/mmol H2O∙m−2∙s−1)、气孔导度(Gs/mol H2O∙m−2∙s−1)、胞间CO2浓度(Ci/μmol CO2∙mol−1)、光合有效辐射(PAR/μmol∙m−2∙s−1)、空气温度(Tair/℃)、大气CO2浓度(Ca/μmol CO2∙mol−1)。水分利用效率(WUE/μmol·CO2∙μmol−1∙H2O)计算公式为WUE = Pn/Tr,气孔限制值(Ls)计算公式为Ls = 1 − Ci/Ca。
所有数据取平均值,用Microsoft Excel 2016软件进行计算和各因子日变化曲线图绘制,并通过SPSS26.0软件对胡杨叶片各光合参数和环境因子的平均值进行方差、相关性、多元逐步回归分析。
3. 结果与分析
3.1. 胡杨雌雄光合参数在各冠层的日变化
净光合速率是表征植物光合作用强弱的一个指标。胡杨雄株净光合速率(Pn)日变化在冠层10 m处呈现“双峰”型,其余冠层均呈现“单峰”型(图1)。胡杨雌雄株的Pn均随冠层高度的增加而升高;在9:30~11:30,随着光照增强,气温增高,胡杨Pn快速升高,在11:30达到最大值为13.64 μmol CO2∙m−2∙s−1;在11:30~17:30,随着辐射减弱,气温逐渐降低,Pn逐渐下降。在15:30,胡杨雄株10 m处的Pn显著高于其他冠层(表2)。胡杨雌株Pn日变化除了冠层6 m呈现出“双峰”型,其余冠层均呈现“单峰”型,且在11:30 Pn达到峰值,冠层10 m处达到最大值,为15.19 μmol CO2∙m−2∙s−1,显著高于其他冠层(表2)。
雌雄株间的Pn比较发现,从9:30~17:30,雌雄株Pn的峰值均出现在11:30,从冠层2 m到10 m雄株比雌株分别高了1.9%、7.9%、8.7%和10.2%;从9:30~11:30,雌雄株各冠层的净光合能力增幅最大,且雄株各冠层的增幅大于雌株;同时还发现雄主雌株10 m冠层处在11:30和15:30出现了双峰现象,分别为13.64 μmol CO2∙m−2∙s−1和10.98 μmol CO2∙m−2∙s−1。
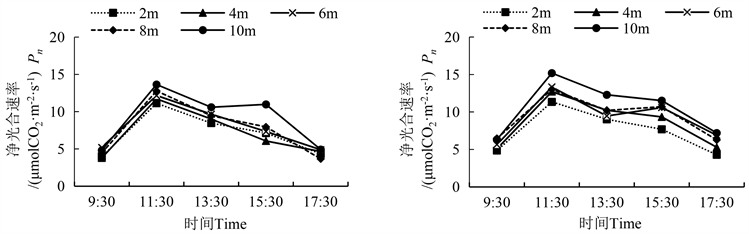
Figure 1. Diurnal variation of net photosynthetic rate of male and female Populus euphratica (left is male, right is female)
图1. 胡杨雌雄净光合速率的日变化 (图左为雄,图右为雌)
气孔导度反映植物叶片气孔与外界进行气体交换的通畅程度。从图2可以看出,胡杨雄株气孔导度(Gs)日变化在冠层6 m、8 m、10 m呈单峰曲线,在冠层4 m则表现出“先上升,下降,再上升”的趋势。胡杨雌株Gs日变化各冠层均呈双峰曲线,于11:30出现最高峰,15:30出现次高峰。在不同时段,胡杨雌雄株Gs日变化于11:30出现最大值,在11:30~15:30呈现上下波动,但变化幅度不大,在15:30~17:30快速下降。在11:30、13:30、15:30处,雌雄株冠层2 m与10 m处Gs均有显著差异;在同一时间,胡杨雌雄株冠层10 m处的Gs日变化幅度较冠层2 m处大(表2)。
雌雄株间的Gs比较发现,从9:30~11:30,冠层4 m、8 m、10 m处雄株的增幅为144.7%、171.7%和102.6%,均大于雌株的增幅,且从11:30~17:30,8 m、10 m冠层处雄株的降幅大于雌株,分别降低了59.5%、71.0%。
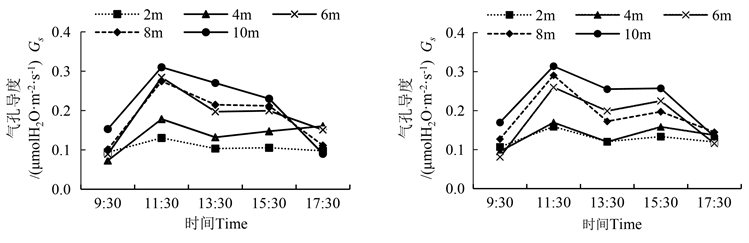
Figure 2. Diurnal variation of male and female stomatal conductance of Populus euphratica (left is male, right is female)
图2. 胡杨雌雄气孔导度的日变化(图左为雄,图右为雌)
Table 2. Comparison of net photosynthetic rate and stomatal conductance of leaves between male and female plants of Populus euphratica in different canopies
表2. 胡杨雌雄株不同冠层间叶片净光合速率、气孔导度比较
注:同一行不同字母表示显著性差异(P < 0.05)。
胡杨雌雄株的胞间CO2浓度(Ci)日变化均呈现“V”字型,在13:30出现最低值(图3)。Ci日变化规律基本与Pn相反,因为当净光合速率较大时,固定的CO2较多,引起了胞间CO2浓度降低。在各冠层高度上,胡杨雌雄株的Ci均随着冠层的升高而减少。同一时间不同冠层之间的Ci值均不显著(表3)。
雌雄株间的Ci比较发现,雌株冠层2 m到10 m处在 9:30~13:30,分别降低了18.7%、13.7%、14.1%、22.5%和24.0%,均大于雄株的降幅,而在13:30~17:30,分别增加了23.1%、17.4%、19.2%、27.9%和30.0%,均大于雄株的增幅。
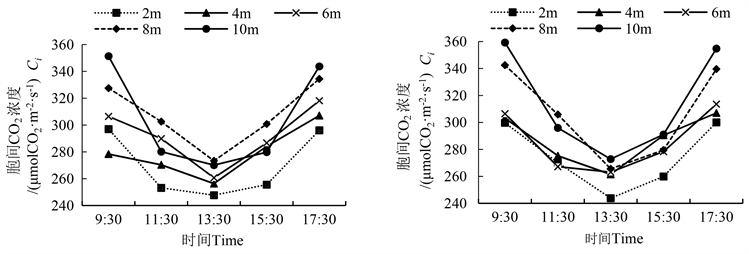
Figure 3. Diurnal variation of CO2 concentration between male and female cells of Populus euphratica (left is male, right is female)
图3. 胡杨雌雄胞间CO2浓度的日变化(图左为雄,图右为雌)
Table 3. Comparison of intercellular CO2 concentration and water use efficiency in male and female leaves of Populus euphratica in different canopies
表3. 不同冠层胡杨雌雄叶片胞间CO2浓度、水分利用效率比较
注:同一行不同字母表示显著性差异(P < 0.05)。
WUE是表征植物光合与蒸腾的竞争能力。胡杨雌雄株各冠层高度的水分利用效率(WUE)均呈上下波动趋势(图4),均在11:30达到最大值,在13:30达最低值,其原因是这一时段温度升高使得胡杨叶片有更大的蒸腾量,气孔导度增大,加大了水分散失,导致水分利用效率降低。不同冠层间比较,冠层2 m的WUE高于冠层8 m、10 m。在11:30、15:30,胡杨雌株冠层2 m与10 m的WUE值差异显著;在11:30、13:30、15:30、17:30,胡杨雄株冠层2 m与10 m的WUE值差异显著(表3)。
雌雄株间的WUE比较发现,在13:30、15:30,雌株冠层10 m处的WUE显著高于雄株(表4)。除9:30,各时间点雄株2 m处冠层的WUE均大于其余各冠层。雌雄株WUE的最小值均出现在13:30,且2 m到10 m处,雄株比雌株分别低0.1%、33.0%、1.9%、23.7%和26.3%;从11:30~13:30,雄株各冠层分别降低了45.4%、60.0%、58.2%、58.9%和48.1%,大于雌株降低的31.8%、41.9%、43.2%、52.1%和39.7%。
图5显示,胡杨雌雄株各冠层的蒸腾速率(Tr)呈现单峰曲线,均在13:30出现最大值,之后呈现下降趋势(图5)。冠层8 m、10 m的蒸腾速率要高于冠层2 m、4 m。冠层10 m的Tr日变化幅度较2 m大,在9:30、13:30、15:30,雌株冠层2 m与8 m、10 m的Tr值差异显著;在17:30,雄株冠层2 m与8 m、10 m的Tr值差异显著(表5)。
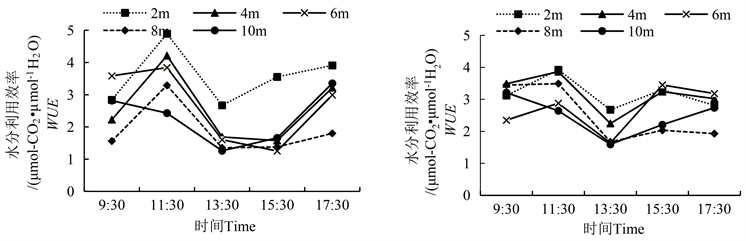
Figure 4. Diurnal variation of water use efficiency between males and females of Populus euphratica (left is male, right is female)
图4. 胡杨雌雄水分利用效率的日变化(图左为雄,图右为雌)
Table 4. Comparison of water use efficiency and transpiration rate of male and female Populus euphratica in the same canopy
表4. 同一冠层胡杨雌雄水分利用效率、蒸腾速率比较
注:雌雄之间不同字母表示显著性差异(P < 0.05)。
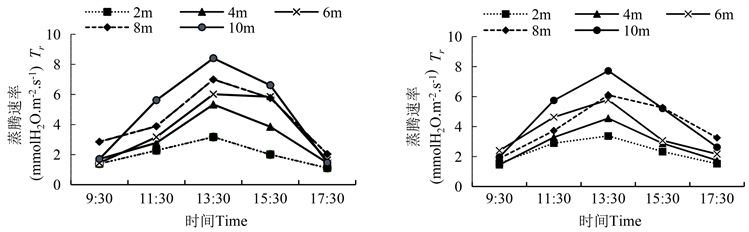
Figure 5. Diurnal variation of male and female transpiration rate of Populus euphratica (left is male, right is female)
图5. 胡杨雌雄蒸腾速率的日变化(图左为雄,图右为雌)
Table 5. Comparison of transpiration rate and stomatal limitation of leaves of Populus euphratica in different canopies
表5. 不同冠层胡杨叶片蒸腾速率、气孔限制值比较
注:同一行不同字母表示显著性差异(P < 0.05)。
雌雄株间的Tr比较发现,在11:30、13:30,冠层4 m、6 m的胡杨雌雄株Tr值差异显著(表4)。雌雄株各冠层的Tr最大值均出现在13:30。从9:30~13:30,雄株在冠层2 m、4 m、6 m、10 m处,分别增加了127.8%、212.7%、315.3%和391.4%,均大于雌株的增幅,而从13:30~17:30,在2 m到10 m处,各冠层分别降低了64.9%、72.8%、72.8%、70.9%和82.5%,降幅均大于雌株。
由图6可知,胡杨雌雄株的气孔限制值(Ls)日变化均呈“单峰”型,于13:30出现峰值,胡杨雄株的Ls最大值为8.41 mmol H2O∙m−2∙s−1,雌株的Ls最大值为7.72 mmol H2O∙m−2∙s−1,之后呈下降趋势。冠层间相比,冠层2 m的Ls高于冠层8 m、10 m,与WUE相一致,雌株的WUE与Ls显著相关,雄株极显著相关(表6),说明气孔限制值影响水分利用效率的大小。
雌雄株间的Ls比较发现,在13:30,雌雄株冠层2 m与10 m的Ls值差异显著(表5)。9:30~13:30,雌株冠层2 m、4 m、8 m、10 m处Ls值增加了53.0%、39.0%、120.1%、185.1%,均大于雄株的增幅,而在13:30~17:30,雌株2 m、8 m、10 m冠层处降低了34.2%、54.2%和62.3%,均大于雄株的降幅。
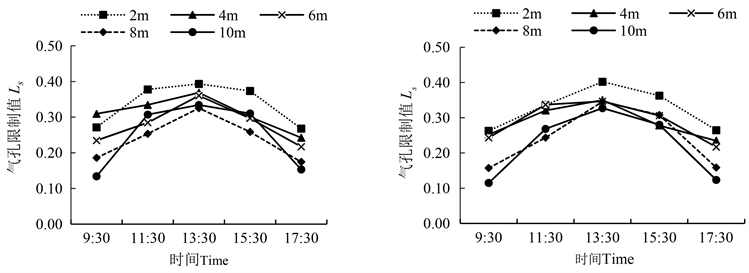
Figure 6. Diurnal variation of male and female stomatal limit of Populus euphratica (left is male, right is female)
图6. 胡杨雌雄气孔限制值的日变化(图左为雄,图右为雌)
3.2. 影响胡杨雌雄株光合特性的内外因素
相关分析可以反映出两个变量之间线性关系强度及方向。对各环境因子、冠层、叶面积和各光合参数的平均值用SPSS26.0进行相关性分析(表6)。结果可知,在各生理因子中,胡杨雌株的Ls与Tr呈极显著正相关(P = 0.309**),而雄株无明显相关性(P = 0.208);胡杨雌株的Ls与WUE呈显著正相关(P = 0.281*),而雄株呈极显著正相关(P = 0.388**);而对于各环境因子,胡杨雌雄株的Tair均与WUE呈极显著负相关;胡杨雌株PAR与WUE无明显相关性(P = −0.166),雄株呈显著负相关(P = −0.262*);胡杨雌株Ca与Pn、Ls均呈显著负相关,与Ci呈显著正相关(相关系数依次为−0.232*、0.315**和−0.318**);而雄株均无明显相关性(相关系数依次为−0.074、0.165、−0.176);胡杨雌株的Pn与冠层之间呈显著正相关(P = 0.289*),而雄株无明显相关性(P = 0.188);胡杨雌株的叶面积La与Tr呈显著正相关(P = 0.273*),而雄株无明显相关性(P = 0.145)。综合来看,胡杨雌株叶片的光合作用受生理因子和环境因子的影响比雄株更显著。
Table 6. Correlation coefficients between photosynthetic parameters of male and female leaves of Populus euphratica and environmental factors, canopy height and leaf area
表6. 胡杨雌雄叶片光合参数与环境因子、冠层高度及叶面积的相关系数
注:*、**分别表示在0.1、0.05水平上的显著。
3.3. 光合参数与环境因子的线性回归分析
回归分析是研究一个变量随另一个变量变化的方法。为深入了解胡杨雌雄株之间净光合速率、蒸腾速率、胞间CO2浓度、气孔导度与环境因子的数量关系,分别将胡杨雌雄的Pn、Tr、Ci、Gs与PAR、大气二氧化碳浓度Ca、Tair 3个环境因子进行逐步回归分析,依次得到最优回归方程(表7),决策系数(Rreg 2)均达到显著水平(P = 0.000)。从回归方程可以看出,对胡杨雌株Pn的促进作用最大的是PAR,而对胡杨雄株Pn的促进作用最大的是Tair;对胡杨雌雄株Tr促进作用最大的均是Tair;对胡杨雌株Ci的抑制作用最大的是PAR,而对胡杨雄株Ci的抑制作用最大的是Tair;对胡杨雌株Gs的促进作用最大的是Tair,3个环境因子均对胡杨雄株Gs无显著影响。PAR是影响胡杨雌株叶片净光合速率最主要的环境因子;Tair是影响胡杨雄株叶片净光合速率最主要的环境因子。
Table 7. Optimal regression equations of photosynthetic parameters and environmental factors in leaves of male and female Populus euphratica
表7. 胡杨雌雄株叶片光合参数与环境因子的最优回归方程
4. 讨论与结论
光合作用是植物有机物质积累、生长发育的源头,受植物本身生理过程和环境因素(太阳辐射、大气 CO2浓度、土壤水分等)的共同影响。不同植物光合作用变化特性受自身生理因子和环境因子的影响而表现出不同的趋势。黄芩(Scutellaria baicalensis Georgi)的光合日变化呈不明显双峰曲线,有轻微光合“午休”现象 [22] ;靳甜甜等(2011)对沙棘的研究中,中、上坡位出现了明显的光合“午休”现象,而下坡位未出现明显午休现象 [23] 。中、上坡位出现光合“午休”现象时,Ci降低而Ls升高,说明Gs降低引起的光合原料短缺和光合产物的积累是引起Pn降低的主要原因;金则新等(2004)曾对自然分布在高山生长的云锦杜鹃进行日变化分析,结果发现阳叶呈现“双峰”型,其阴叶却呈现“单峰”型,说明叶片光合“午休”由叶片所处的环境因素决定,并非其固有的生理现象 [24] ;常宗强等(2006)测定了额济纳洲胡杨光合速率日变化,并通过控制光照强度和CO2浓度研究Pn对其的响应特征,结果表明Pn随着PAR和Ca增大而增大 [18] ;庞发虎等(2010)模拟了不同干旱程度,对杨树生理生态因子与环境因子关系进行研究,杨树生长受到PAR、Ca、Tair等的显著影响,且生理变化宜综合多个指标共同分析 [25] 。代雅琦等(2021)研究发现胡杨光合速率日变化呈“单峰”型,这与本研究结果一致 [16] 。本研究结果发现,胡杨雌雄株Pn、Gs、Tr、Ls的日变化均是先增加后减小的趋势,而Ci呈现先减小后增加的趋势,且Pn均与叶面积、环境因子(PAR、Tair、Ca)呈显著正相关,这与前人研究结果一致 [26] 。中午Pn降低的原因可能是因为温度升高引起气孔导度降低,或是PAR迅速升高超过光饱和点而致使Pn下降。
树冠作为树木的主要组成部分,它与树木的光合作用、呼吸作用、蒸腾作用等一系列生理活动都有密切的联系,不仅体现了树木进行光合作用的面积和有机物传输的方向及距离,还反映了光合作用的效率,其大小对树木的生长活力和生产力状况起着决定性的作用,同时不同冠层位置叶片的光合能力存在差异 [27] 。本研究结果表明,胡杨位于冠层10 m叶片的光合能力显著高于冠层2 m。Schulze等(1977)在对挪威云杉(Piceaabies)冠层光合作用的研究中得到类似的结论 [28] 。Kull (2002)表明下层光合产量偏低的主要原因是光照条件不足 [29] 。本研究结果发现,从9:30~11:30,Pn、Gs、Tr的增幅雄株大于雌株,但在13:30,胡杨雌株的净光合速率、气孔导度高于雄株,雌株冠层10 m的WUE显著高于雄株。9:30~13:30,雌株冠层2 m、4 m、8 m、10 m处Ls值的增幅大于雄株,而在13:30~17:30,雌株的降幅同样大于雄株。说明在正午温度升高,干旱胁迫加重的情况下,雄株气体交换能力变化较平缓,说明雄株本身被干旱环境影响较小,在干旱环境下更稳定。胡杨雌株叶片的光合作用受生理因子和环境因子的影响比雄株更显著。
对Pn、Tr、Ci、Gs与PAR、Ca、Tair 3个环境因子进行逐步回归分析,结果表明PAR是影响胡杨雌株叶片净光合速率最主要的环境因子,这与众多学者的结果相同 [26] [30] [31] ;而Tair是影响胡杨雄株叶片净光合速率最主要的环境因子,Tair对胡杨Pn表现出了促进作用,与吴桂林等(2016)的研究结果相同 [32] ,而对雌雄株Tr影响最大的均是Tair。综上所述,胡杨雌雄株光合日变化受生理因子和环境因子共同作用,但对净光合速率影响的环境因子在性别间不同,这可能是性别间光合生理内部机理的不同。
基金项目
感谢国家自然科学基金项目(U1803231, 31860198)的资助。
NOTES
*通讯作者。