摘要: 本研究建立一种新的生态调控模式,通过在有河蟹的稻田围隔内加入含有乳酸菌,反硝化细菌、芽孢杆菌为主的复合微生态制剂,探讨微生态制剂对养殖水体中溶解氧,pH,氨态氮、亚硝酸氮和活性磷以及河蟹生长和水稻产量的影响。通过研究得出以下结论:在稻蟹种养田水体内添加微生态制剂可以显著影响水质,与对照组进行比较,试验组的pH、溶解氧、氨态氮、亚硝酸氮和活性磷有显著性差异(P < 0.05)。其中溶解氧降低14%,氨态氮升高53%,亚硝酸氮升高50%,活性磷升高20%。但氨态氮,亚硝酸氮和活性磷相对含量都很低,同时也为水体带来了N和P等营养物质。水体细菌群落没有显著变化,优势门为变形菌门、拟杆菌门、放线菌门、厚壁菌门。优势属分别为Limnohabitans、Hydrogenophaga、多核杆菌属、黄杆菌属,Curvibacter。水稻千粒重增加9.14%,扣蟹质量增加11.87%,且均没有病害发生。因此,在稻蟹种养田中应用微生态制剂对水质有显著影响,水体细菌群落基本保持稳定,水稻和扣蟹产量上升,有较好的经济增加产出。
Abstract:
In this study, a new eco-logical regulation model was established, and the effects of microecological preparations on dis-solved oxygen, pH, ammonia nitrogen, nitrite nitrogen and active phosphorus, as well as Eriocheir sinensis growth and rice yield were explored by adding compound microecological preparations mainly containing lactic acid bacteria, denitrifying bacteria and bacillus in paddy fields with Eri-ocheir sinensis. Through the study, it was concluded that the addition of microecological prepara-tions to the water body of rice crab cultivation field could significantly affect the water quality, and compared with the control group, there were significant differences in pH, dissolved oxygen, ammo-nia nitrogen, nitrite nitrogen and active phosphorus in the experimental group (P < 0.05). Among them, dissolved oxygen decreased by 14%, ammonia nitrogen increased by 53%, nitrite nitrogen increased by 50%, and active phosphorus increased by 20%. However, ammonia nitrogen, nitrite nitrogen and active phosphorus are relatively low in content, and they also bring nutrients such as N and P to the water body. There was no significant change in the bacterial community of the water body, and the dominant phylum were Proteobacteria, Bacteroides, Actinomycetes, and Firmicutes. The dominant genera are Limnohabitans, Hydrogenophaga, Polynucleobacteria, Flavobacteria, and Curvibacter. The weight of 1,000 grains of rice increased by 9.14%, and the mass of buckle crab in-creased by 11.87%, and no diseases occurred. Therefore, the application of microecological prepa-rations in rice crab breeding fields had a significant impact on water quality, the bacterial commu-nity of the water body remained basically stable, the yield of rice and juvenile crabs increased, and there was a good economic increase in output.
1. 引言
稻蟹共生是一种立体生态农业种养新模式,有利于稻和蟹双方的物质和能量进行良性循环流动,从而增加收入和资源利用的功能体系。推广稻田养殖技术,既能很好利用农村资源、提高土壤肥力和增加农民收入,又可以转移部分农村剩余劳动力,还能提高农民科学技术水平和生产技能 [1] 。一方面,水稻为河蟹提供了一个有利于生存的环境,另一方面,河蟹为水稻提供了肥料,促进生长,减少了田间病害的发生率,稻田生态养蟹的水质动态变化对水稻和河蟹生长均存在着至关重要的影响 [2] 。
水质因子包括溶解氧、pH、水温、三态氮、磷、钾等。各因子间相互作用、相互影响、相互转化,一个因子的变化可能是由某一个因子引起的,可能是由多个因子引起的,其中的每一个因子都能对稻田水环境造成影响 [3] 。
微生态制剂是从自然环境中筛选得到微生物,再经过培养、繁殖之后,制成包含大量微生物的活菌制剂,是新兴的养殖添加剂,又被称为微生态调节剂。对于水产养殖来说,微生态制剂遵从微生态理论的指导,治理水污染环境,维持水生态平衡,而且能预防病原菌对水产养殖生物的入侵,提高水产养殖生物的健康水平及产量 [4] 。对于水体环境改善来说,微生态制剂具有无残留、不会造成二次污染,无毒且无副作用,不仅可以用来净化水质、改善水生态环境,而且还能有效防治病害、增强生物的免疫力、提高人工饲料的生物利用率和养殖生物生长繁殖性能,降低养殖水体及粪便中氨、磷等有害物质的含量,减少氨态氮、磷等对水体的污染,更重要的是减少养殖水体对周边环境所造成的污染 [4] [5] [6] 。微生态制剂的原理是利用菌群之间存在着相互竞争的机制,向水体中定量的投加能够改良水质、分解有机物和有毒物质,且对养殖生物不会产生危害、对水体无二次污染的生物制剂,将会在水体中建立一种新的动态平衡,使有毒物质得以分解,有害病原得以抑制,水体水质得到改善,水产养殖生物可以健康生长 [7] 。目前,国内外研究的微生态制剂包括单一菌种和复合菌种,单一菌种主要有枯草芽孢杆菌;复合菌种为多种有益菌按比例搭配而成 [8] [9] 。
微生态制剂的应用可以改善稻田水体水质 [10] [11] [12] ,促进河蟹生长 [13] [14] ,还可提高水稻产量 [15] [16] [17] [18] ,谷加坤等 [19] 研究证明了微生态制剂可以增加养蟹水体微生物群落多样性,但对于稻田水体的研究较少,为此,本研究中探讨了三种微生态制剂对稻田水质、水体细菌群落、水稻生长及河蟹产量的影响,为微生态制剂在稻蟹种养田的广泛和高质量应用提供参考。
2. 材料和方法
2.1. 试验地点
试验在盘锦光合蟹业有限公司稻田进行。位于辽宁盘锦大洼地区(N 40˚90';E 121˚85')。年均气温8.5℃,平均降水量650 mm,无霜期180天,为一年一季水稻种植区。
2.2. 试验材料
水稻品种为盐粳927,大眼幼体来源于盘锦光合蟹业有限公司培育的蟹苗。微生态制剂为配制的三种复合微生态制剂。水嫩爽(酵母菌、乳酸菌、芽孢杆菌等)活菌量为5.0 × 105 cfu/g。黑变黄(酵母菌、乳酸菌、芽孢杆菌、硝化细菌、反硝化细菌)活菌量为5.0 × 105 cfu/g。高抗一号(酵母菌、乳酸菌等)活菌量为1.0 × 105 cfu/g。
2.3. 试验设计
本试验选取2个相同规格地块,试验分为两组,A为对照组,B为试验组。每组一个地块,每个地块内有相邻的8个围隔,选取其中间的3个围隔为每个分组的平行,每个围隔面积为5 × 5 m2,水深6~20 cm,围隔内有水稻,投放大眼幼体约2500只。每个小组在整个试验周期内,不加肥不喂料。对照组不加入微生态制剂,试验组于8月11日、8月17日、8月25日、9月2日、9月14日、分别加入微生态制剂,每个地块10 kg水嫩爽,10 kg黑变黄,5 kg高抗一号均匀泼洒到8个围隔里,当天天气状况均为良好,整个试验时长为41 d。
2.4. 采样与分析方法
在整个试验周期内,取每块地中间的三个围隔的样本,每个稻田围隔内设4个取样点(a、b、c、d),分别位于围隔四个角即东南(a)、东北(b)、西北(c)、西南(d),每个取样点距稻田围隔侧边各15 cm左右,每天每个监测点分别取表层水样各一瓶混合,采样时间为16:00,当天天气均为良好。测定加菌与不加菌稻田水质的水平变化。
所测的水环境指标包括:溶解氧(DO)、pH、氨态氮(NH4-N)、亚硝酸氮(NO2-N)和磷酸盐(PO4-P)。所有水样经分类编号后,在公司水化学试验室2 h内完成测定。DO和T采用美国YSI550A便携式溶解氧测定仪直接测定,pH采用便携式pH计(PHB-1,上海三信仪表厂)测量,S用便携式盐度计测定,NH4-N含量采用纳氏试剂分光光度法测定;NO2-N采用N-(1-萘基)-乙二胺光度法;PO4-P采用钼酸铵分光光度法测定。
试验结束时将各组水样由真空泵经微孔滤膜50 mm/0.45μm (直径/孔径)进行抽滤,收集菌体,将所抽好的滤膜放在无菌离心管中,保存于−80℃的超低温冰箱,由北京百迈克生物科技有限公司进行DNA提取与高通量测序。
2.5. 水样DNA提取、PCR扩增和建库测序流程
无菌条件下将−80℃保存的滤膜研磨称重后进行水样DNA的提取。使用TGuide S96磁珠法土壤/粪便基因组DNA提取核酸试剂盒提取总DNA,使用16S全长序列引物27F和1492R (Lane, 1991)。PCR扩增在总体积为30 uL的溶液中进行,其中含有1.5 uL基因组DNA、10.5 μL NFW、15 uL KOD ONE MM、3 uL barcode 引物对。PCR反应条件为95℃ 2 min,98℃ 10 s,55℃ 30 s,72℃ 1.5 min,72℃ 2 min,25个循环 [7] 。
将PCR产物纯化、定量并均质化以形成测序文库(SMRT-Bell)。在对文库进行量化后,使用单分子测序仪(PacBio SequelII)进行高通量测序。机外数据采用bam文件格式,CCS (循环共识排序)文件由smrtlink分析软件(SMRTLink,8.0版)导出。根据序列识别来自不同样品的数据,并将其转换为fastq数据集。运用ylima(vl.7.0)软件识别不同样品的CCS序列,并去除嵌合体,从而生成高质量CCS序列 [8] 。
2.6. 数据处理与分析
OTU即分类操作单元,是在系统发生学研究或群体遗传学研究中,为了便于进行分析,人为给某一个分类单元(品系,种,属,分组等)设置的同一标志。可以根据不同的相似度水平,对所有序列进行OTU划分,每个OTU对应于一种代表序列。使用Usearch软件对Reads在97 %的相似度水平下进行聚类、获得OTU。通过RDF分类器(v2.2)使用SILVA数据库(release132)对OTU进行分类注释(置信阈值为80%)。再使用PyNAST (V1.2.2)将选定的OTU与SILVA数据库的核心比对模板进行比对。以显示细菌群落组成的差异。使用QIIME2软件,对α多样性指数进行评估,比较不同样品在物种多样性方面存在的相似程度。
基于Beta这种多样性的分析重建方法得到的四种距离矩阵,通过使用R语言的聚类工具可以采用非加权组物种配对平均法(UPGMA)对每一个样品间物种进行系统的层次聚类,以准确地判断各个物种的样品间相似度和物种层次发生组成的系统相似性。其原理是:假定的前条件是在进化过程中,每一世系发生趋异的次数相同,即核苷酸或氨基酸的替换速率是均等且恒定的。通过UPGMA法所产生的系统发生树可以说是物种树的简单体现,在每一次趋异发生后,从共同祖先节点到2个特征间的支的长度一样。
试验数据均以平均值 ± 标准差(mean ± S.D.)表示,采用SPSS 26.0软件对试验数据采用独立样本t检验比较,以P < 0.05为差异显著水平,P < 0.01为差异极显著水平。
3. 结果与分析
3.1. 微生态制剂对稻田水质的影响
由图1可知,试验期间,B组pH变化范围为(7.19~8.19),平均为(7.62 ± 0.20)。A组pH变化范围为(7.16~7.83),平均为(7.43 ± 0.13) mg/L,试验组与对照组有显著差异(P < 0.05)。
由图2可知,试验期间,DO含量变化较大,B组变化范围为(0.17~5.58) mg/L,平均为(2.67 ± 1.37) mg/L。A组变化范围为(1.99~4.25) mg/L,平均为(3.11 ± 0.56) mg/L,对照组与试验组差异显著(P < 0.05),加入微生态制剂之后,DO含量明显降低,此外,还受天气的影响,晴天时含量升高,阴天下雨时含量降低。
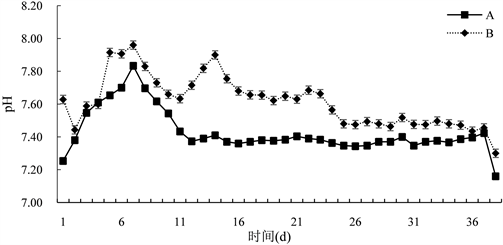
Figure 1. Changes in pH of paddy water bodies
图1. 稻田水体pH的变化
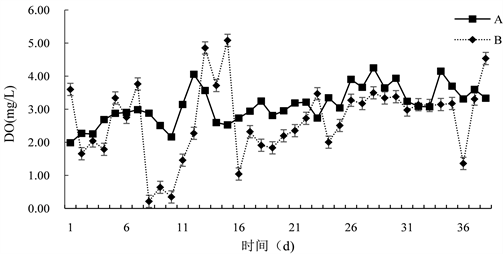
Figure 2. Changes of dissolved oxygen content in water of paddy field
图2. 对稻田水体溶解氧含量的变化
由图3可知,试验期间,B组NH4-N变化范围为(0.80~2.57) mg/L,平均为(1.35 ± 0.41) mg/L,A组NH4-N变化范围为(0.60~1.23) mg/L,平均为(0.82 ± 0.15) mg/L,对照组与试验组差异极显著(P < 0.01);每次加入微生态制剂之后,试验组NH4-N含量明显升高,之后随着加入次数的增多,NH4-N含量升高的趋势越不明显,试验中前期较中后期波动明显,对照组试验期间变化不明显。
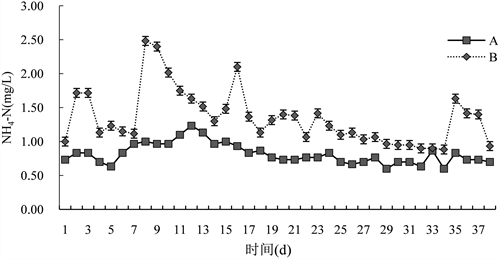
Figure 3. Changes in ammoniacal nitrogen levels in paddy water bodies
图3. 对稻田水体氨态氮含量的变化
由图4可知,试验期间,B组NO2-N变化范围为(0~0.07) mg/L,平均为(0.03 ± 0.01) mg/L,A组NO2-N变化范围为(0.01~0.03) mg/L,平均为(0.02 ± 0.01) mg/L,对照组与试验组差异极显著(P < 0.01);但各组NO2-N含量很低,且没有明显升高,18 d前基本保持在0.02 mg/L以下,与对照组差异不显著,试验中后期试验组NO2-N含量呈略微上升趋势,第21 d NO2-N含量最高,为0.07 mg/L。
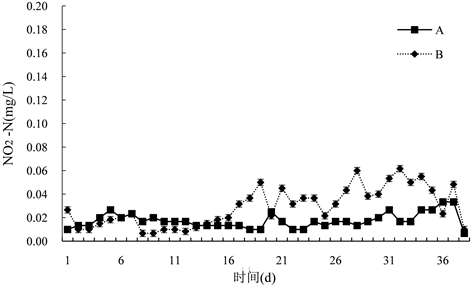
Figure 4. Changes of water nitrite content in paddy field
图4. 对稻田水体亚硝酸氮含量的变化
由图5可知,试验期间,B组PO4-P变化范围为(0.06~0.34) mg/L,平均为(0.12 ± 0.06) mg/L,A组PO4-P变化范围为(0.05~0.16) mg/L,平均为(0.10 ± 0.01) mg/L,对照组与试验组差异显著(P < 0.05)。前期波动较大,试验组PO4-P含量呈先上升后下降趋势,15 d后,对照组与试验组PO4-P含量基本保持不变且无明显变化趋势。
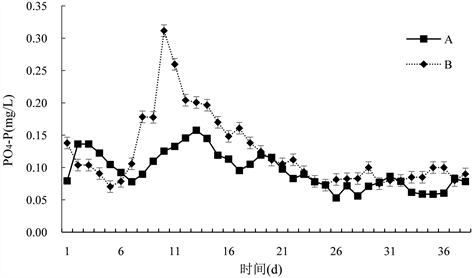
Figure 5. Changes in the content of active phosphorus in paddy water
图5. 对稻田水体活性磷含量的变化
3.2. 微生态制剂对稻田水体细菌群落组成的影响
3.2.1. Alpha多样性指数
细菌群落的Alpha多样性指数如表1所示,试验组与对照组的OTU数目、物种多样性指数(Shannon, Simpson)和物种丰富度指数(Ace, chao1)均无显著差异(P > 0.05)。

Table 1. Alpha diversity index statistics
表1. Alpha多样性指数统计
3.2.2. Venn图
Venn图(图6)显示水体中OTU总数为1099个,其中对照组A与试验组B共有OTU为712个,A组特有OTU为98个,占总OTU的8.92%;B组特有OTU为289个,占总OTU的26.30%。
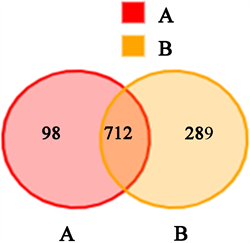
Figure 6. Venn diagram of water body in paddy field
图6. 稻田水体Venn图
3.2.3. UPGMA分析
UPGMA (Unweighted Pair-Group Method with Arithmetic Mean)即非加权组平均法,以判断各样品间物种组成的相似性。如图7,从树状关系形式可视化分析的结果可知,可以将其分为两大部分第一部分主要是A1和B1,末端竖线相聚在一起,相似度较高。第二部分主要包括A4、A7、B7、B4,末端竖线相聚在一起,相似度较高,其中A4和A7相似度最高。由此可知B组的平行样本之间相似度比A组大。
3.2.4. 稻田水体门水平细菌群落组成
试验组与对照组样品细菌群落组成前10的门分别为变形菌门、拟杆菌门、放线菌门、厚壁菌门、Campilobacterota、疣微菌门、蓝细菌门、Epsilonbacteraeota、螺旋体门、Patescibacteria (图8)。其中变形菌门、拟杆菌门、放线菌门、厚壁菌门为优势门(相对丰度 > 1%)。试验组与对照组中四个门占比依次为58.50%和51.43%、17.72%和22.22%、15.10%和21.99%以及2.61%和1.03% (图8)。与对照组相比,试验组变形菌门和厚壁菌门丰度增加,放线菌门和拟杆菌门丰度降低,但t检验结果表明10个门差异不显著(P > 0.05)。
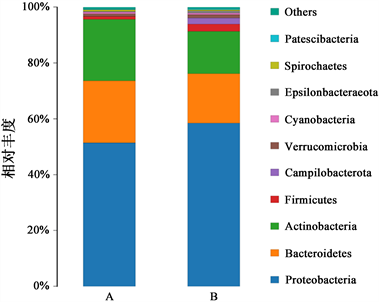
Figure 8. The bacterial community compositionat thephylum level. Note: only the phyla in the top 10 of the abundance level are shown, and the other phyla are merged into others shown in fig.8
图8. 门水平群落组成图。注:只显示丰度水平前10 的门,并将其他物种合并为others在图中显示
3.2.5. 稻田水体属水平细菌群落组成
试验组与对照组样品细菌群落组成前10的属分别为Limnohabitans、Hydrogenophaga;多核杆菌属、黄杆菌属,Curvibacter,Aurantimicrobium,Rhodocluna,Allopseudarcicella,Candidatus_Planktoluna和Candidatus_Planktophila (图9)。2个组的水体细菌群落组成均以Limnohabitans、多核杆菌属、Curvibacter和Rhodocluna为主。
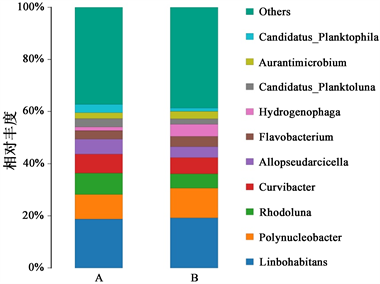
Figure 9. The bacterial community composition at the genuslevel. Note: only the genera in the top 10 of the abundance level are shown, and the other genera are merged into Others shown in fig
图9. 属水平细菌群落组成表。注:只显示丰度水平前10的属,并将其他物种合并为Others在图中显示
相比于对照组,试验组的Limnohabitans、Hydrogenophaga、Flavobacterium、多核杆菌属丰度升高;Curvibacter、Rhodocluna和Allopseudarcicella丰度下降。通过t检验得出10个属差异不显著(P > 0.05)。
3.3. 水稻和扣蟹产量
最后经烘干测产(表2),B组水稻千粒重平均为26.99 ± 0.70 g,A组平均千粒重为24.73 ± 0.24 g,两者差异极显著(P < 0.01)。B组扣蟹单位重量为4.63 ± 0.69 g,A组扣蟹单位重量为5.18 ± 0.94 g,两者差异不显著(P > 0.05)。
Table 2. Physiological morphology of rice and yield of juvenile crab in each group
表2. 各组水稻的生理形态和扣蟹产量
注:同列中标不同字母者表示组间有显著性差异(P < 0.05),标相同字母者表示组间无显著性差异(P > 0.05)。
Note: Those marked with different lowercase letters in the same column indicated significant difference between groups (P < 0.05), those marked with the same lowercase letters indicated no significant difference between groups (P > 0.05).
4. 讨论
4.1. 微生态制剂对稻田水质的影响
最适宜河蟹生长的水质为中性至弱碱性,pH为7.5~8.5,试验期间的稻田水体基本符合范围,微生态制剂对于pH的影响不大,影响pH的主要因素为天气,阴天下雨时pH降低,因为雨水呈中性至弱酸性,加上水中藻类光合作用减弱,而且生物的呼吸作用消耗了水中的DO,间接降低了pH。
试验期间DO含量起伏较大,因为微生态制剂含有枯草芽孢杆菌等好氧菌,在加微生态制剂前期,DO水平达最低,后期由于水中藻类的光合作用,DO再次上升。因为河蟹的呼吸机制与鱼类不同,其在陆上和水中都可以进行呼吸作用。因此,河蟹会趋利避害,扣蟹仍然会在安全生长水平以上。由于稻田不同于池塘的环境特点,稻田水浅且水量少,水中DO的补充较快,可以将有机物矿化直至完全氧化,削弱了环境对河蟹的不良影响,因此,加入微生态制剂不会制约河蟹的生长。
芽孢杆菌是一类具有高活性的消化酶系、耐高温、抗应激的异养细菌,能发挥氧化、氨化、硝化、反硝化、解硫、硫化、固氮等作用 [20] 。例如地衣芽孢杆菌能在养殖水体水质条件下降解蛋白质与淀粉,自身也具有良好的脱氮去磷效果。枯草芽孢杆菌和巨大芽孢杆菌具有明显去除氨态氮和亚硝酸氮的功能,在养殖水体中使用芽孢杆菌能有效降低水体中化学需氧量、氨态氮和亚硝酸氮等有害物质,并将其转化为二氧化碳、硝酸盐和磷酸盐等,从而为大量藻类生长繁殖提供其必需的营养,而这些藻类又能通过光合作用为水体提供氧气,从而形成一个良好的生态循环。作用机理为,当芽孢杆菌接种到水体后,依靠自身繁殖吸收水体中的有害物质,净化水质;同时占优势的微生物菌群能竞争性抑制并代替病原菌,切断病原菌的营养源,从而减少或阻止了有害病原体的感染 [21] 。
反硝化细菌有翻底的作用,将底泥中的氮翻出,并利用水体中的有机氮源,氨化反应会使水体中的氨态氮量增加,使氨态氮短期内上升较快。之后氨态氮被乳酸菌和芽孢杆菌等转变为菌体蛋白,无机氮被异养细菌利用,水体中的氨态氮被消耗,后期氨态氮下降。而亚硝酸氮自始至终维持在极低的水平,这与稻田水体有机质含量低,经常性换水和水稻自身吸收净化氮元素相关。试验组活性磷前期略微升高,是微生态制剂带来外源性的磷,水稻没有及时吸收所致 [18] 。
4.2. 微生态制剂对稻田水体细菌群落的影响
生态系统的物质循环能力和能量流动效率不仅与微生物的多样性有关,也与其物种组成和相对丰度相关。研究指出,稻蟹共作模式的水体微生物群落组成均以变形菌门、放线菌门和拟杆菌门为主 [19] 。
各组中变形菌门占比最高,与对照组相比,试验组变形菌门相对丰度增加,放线菌门和拟杆菌门丰度降低,另外,变形菌门能改善水体,抑制腐败菌等病原微生物的生长繁殖,转化N、P等元素,与水体N、P元素紧密相关,与有机质降解和纤维素利用相关,在生态系统的碳循环中起重要作用。此外,试验组中厚壁菌门丰度更高,因为加入的微生态制剂中含有属于厚壁菌门的芽孢杆菌,乳酸菌。研究表明,复合芽孢杆菌可以快速改善水体中N或P,乳酸杆菌能产生乳酸、抗菌肽等物质,能够抑制病原菌的生长,且能在水体中直接或间接分解有机质和转化亚硝酸氮等物质,包括了胞内酶作用、代谢产物乳酸的直接化学反应作用和间接促进微生物反硝化作用3种作用机理,其在养殖水体中主要以间接作用方式发挥作用 [21] 。从属水平上的群落组成来看,2个分组的水体细菌群落组成没有显著,均以Limnohabitans、多核杆菌属、Curvibacter和Rhodocluna为主,相比于对照组,试验组的多核杆菌属丰度升高;Curvibacter、Rhodocluna和Allopseudarcicella丰度下降。拟杆菌门中的黄杆菌属具有反硝化功能,在低氧环境下以硝酸盐作为受体进行无氧呼吸,黄杆菌属在生活污水处理、湿地处理海水养殖废水、生物膜反应器中氮的去除中大量存在或是主要类群 [18] 。在本试验中,试验组水体中的黄杆菌属丰度比对照组略高,这可能与试验组水体中DO含量比对照组低相关。有研究表明 [21] 红杆菌科(Rhodobacteraceae)已在多种不同的污水处理工艺中被发现,并鉴定为对含氮污染物的主要去除菌群、Rhodoluna为红杆菌科,本试验中该属的细菌在对照组和试验组中均存在,说明稻田水体中有自我去除含氮污染物的调节能力。
本试验中,加入复合微生态制剂后的稻田水体细菌群落组成没有显著变化,说明稻蟹共作模式下的水体细菌群落组成基本稳定,形成的小生态系统对碳、氮等营养物质的固定及利用能力很强。
4.3. 水稻和扣蟹产量
稻田水体中氮磷含量间接反映了水稻田的供肥能力。整个水稻生长时期,氮磷指标先高后低,因为开始施肥及加入微生态制剂后,水中氮、磷含量升高,水稻对其吸收慢,大部分溶解于水中,造成水体所测各项水质指标升高。随着水稻生长,水稻对于氮、磷吸收量逐渐增加,一部分氮磷在换水的过程中流失,水体氮磷含量随之逐渐减少。王昂等 [18] 的研究结果也表明,分蘖期和拔节期的水稻对氮素的需求量大,硝酸盐,磷酸盐含量呈现较快的下降趋势,成熟期停止对氮素的吸收。试验组氮磷指标较对照组稍高,且比对照组水稻产量显著提升,说明微生态制剂提供了额外的可供水稻吸收的营养成分。此外,稻蟹共作生态环境中蟹类的排泄物以及投喂的饵料也为水稻提供了营养成分 [22] [23] 。
养蟹田水质管理的好坏直接影响河蟹的生长,加入微生态制剂相当于施肥,水体NH4-N等有害物质指标前期升高较快,中后期氮磷基本保持在一定水平,可能是因为菌藻达到了新的平衡,因此为了不影响河蟹生长,可以考虑暂养晚放 [24] 。蟹在生长过程中会产生代谢产物,比如粪便和残饵以及有机碎屑,而这些物质要靠分解者菌来进行分解,菌把这些有机碎屑、残饵、粪便分解成营养物质转化在水里,可以充当水里面的生物肥,进一步循环在水体中,供藻类、水稻作为营养肥料 [25] 。因此,定期施加微生态制剂有助于水稻和扣蟹的生长。
从经济角度上看,稻田养蟹本就提供了额外的经济产值,对照组扣蟹单位重量为4.63 ± 0.69 g,试验组扣蟹单位产量为5.18 ± 0.94 g,相较于不加微生态制剂的养蟹田,试验组每块稻地围隔增加扣蟹增产11.87%,重约为0.34 kg,经济效益为120元。此外,水稻产量两者千粒重差异显著,而发酵微生态制剂的成本远低于增加的产值,因此,微生态制剂带来的直接经济效益是很可观的 [26] 。
5. 结论
1) 复合微生态制剂可以显著影响稻田水质,提高其氨态氮,降低溶解氧含量。
2) 复合微生态制剂对稻田水体细菌群落组成没有显著影响。
3) 复合微生态制剂可以显著提高扣蟹质量和水稻千粒重,能带来直接的经济效益。
参考文献
NOTES
*通讯作者。