1. 引言
外来植物入侵对全球和区域生物多样性和生态系统构危害甚巨,而这些外来植物中有相当一部分是是克隆植物 [1] [2] [3]。有调查显示,我国的外来入侵植物有近半数(44%)为克隆植物,且恶性克隆杂草占据很大比例 [3]。前人研究发现入侵性外来克隆植物主要得益于其与非克隆植物截然不同的生活史性状,而克隆整合及其介导的克隆内劳动分工是最为关键的克隆形状 [4] [5] [6]。
克隆整合(又称克隆内生理整合)是克隆植物所特有的属性,是指其相互连接的分株之间可通过连接子(如匍匐茎和根状茎等)传输水分、营养和同化物质等 [4] [6]。在自然界普遍存在的资源异质性生境中,克隆植物互相连接的分株可通过连接子的作用(即克隆整合)发育分工,即以“趋富特化(Specialization for abundance)”的资源分配方式,专化吸收较为充足资源并通过生理整合输送到资源相对贫乏的相连分株,从而使整个基株系统的生物量和适合度也获得增益 [7] [8]。虽然克隆特性诸如克隆整合和劳动分工对外来植物入侵性的影响和作用已广受国内外学者的关注 [4] [8] [9] [10],但不同生态型的入侵植物克隆特性研究却鲜有记载。
喜旱莲子草(Alternanthera philoxeroides (Mart.) Griseb),俗名也称空心莲子草、水花生、革命草等,原产南美洲巴西,为苋科莲子草属多年生草本,水陆两栖,匍匐生长,现遍及我国温带和亚热带地区,广泛分布于农田、河流、沟渠、林地边缘等陆生和水生环境 [11] [12] [13]。因其环境适应性极强,故危害甚重,已被列为环保部首批公开的16种恶性入侵物种之一 [12]。近期研究发现克隆属性诸如克隆整合和劳动分工是喜旱莲子草建群、扩散和入侵的重要原因之一 [9] [10] [14] [15] [16]。鉴于其水陆两栖的生活史性状,故选取两种不同生态型(水生和陆生)喜旱莲子草植株,通过设置光照和土壤营养两种资源交互的异质性环境,研究不同生态型喜旱莲子草的克隆性状及其作用是否存在差异。具体验证以下假设:1) 异质性生境下,喜旱莲子草陆生型和水生型克隆片段均能表现出劳动分工能力;2) 鉴于陆生环境的异质性相较于水生环境更为普遍 [13] [17],我们预测,相比水生型,陆生型喜旱莲子草克隆片段分株之间会有更强的克隆整合能力,更强的劳动分工强度,从而使其生长和克隆繁殖能更受益于匍匐茎连接(克隆整合)。
2. 材料与方法
2.1. 植物的预培养
本实验在江苏江达生态环境科技有限公司温室大棚中进行。2018年5月,在江苏省无锡市太湖贡湖湾周边区域中自然分布的陆生型和水生型喜旱莲子草种群中分别随机挑选长势良好、大小一致且无虫害的50个喜旱莲子草克隆片段作为实验材料。其中每个喜旱莲子草克隆片段均包含四个分株,分别为远离顶端的两个较老分株(称为基部分株),和靠近顶端的两个较嫩分株(称为端部分株),每个克隆片段都取同样位置以保证分株发育阶段相同。随后将挑选的克隆片段随机匍匐种植在塑料培养框(长25 cm × 宽16 cm × 高8 cm)中进行适应性培养,基质为进口营养土(pH 7.03,总有机质含量为7.36 g/Kg,总氮含量为0.34 g/Kg,总磷含量为0.62 g/Kg),每天适量浇水以促进克隆片段生根。在进行一周的预培养,待克隆片段生根后,从水生和陆生型片段中分别选择长势良好大小一致的32个克隆片段进行后续实验。
2.2. 实验设计
本研究采用两因素析因设计(Two-way factorial design),两个因素分别为克隆整合处理(分株之间的匍匐茎保持连接或者切断)和两种生态型(陆生型和水生型),一共四个处理。实验采用匍匐茎切断法(Stolon severing approach)来检验克隆整合作用,即切断连接克隆植物分株之间的匍匐茎来彻底阻断分株间的资源传输作为无克隆整合的对照,匍匐茎保持连接的处理则为有资源传输的克隆整合处理。该处理方法的优势在于可充分评估克隆整合作用发生的程度,且操作简单易行,在克隆整合相关研究中广泛运用。
将两种生态型的克隆片段两端分别匍匐种植于负相关的斑块资源中,其中将基部分株置于高营养低光照斑块,而将端部分株置于低营养高光照斑块。每一克隆片段株种植在两个规格为上底12 cm、下底8.8 cm、高18.8 cm的培养盆中,每一盆放置两个分株。培养基质为3:1体积的洗净的河沙和绿沸石均匀混合物。温室中的自然光照为高光照处理;低光照处理则采用六针织的遮阴网遮蔽自然光的90% (即光照强度是温室内自然光照的10%)。高营养处理与低营养处理为分别添加2 g和0.4 g进口缓释肥(营养成分为N:P:K,16:9:12),并与基质混合均匀。每个处理重复8次。实验期间,温室温度控制在28~35℃,温室内光照为90%自然光照,实验为期8周,待喜旱莲子草繁殖长满培养盆后收获。
2.3. 数据收集
实验收获前,分别统计端株和基株的叶片数和分株数,并用毫米刻度尺测量匍匐茎长。实验收获后将端株和基株所产生的植株分开测量,将所有植株分成根、茎和叶三部分,分别标记后置于75℃烘箱中烘干48 h至恒重后称重,随后分别统计两端分株和整个克隆片段的总生物量、总分株数、总匍匐茎长和根冠比(Root to shoot ratio)等参数以便进行下一步统计分析。
2.4. 数据分析
所有观测的数据均采用SPSS18.0软件来进行统计分析。分析数据之前,先检验数据是否符合正态分布和方差齐次性。若数据不齐,则进行必要的转换后再进行分析。利用Two-way ANOVA方差分析法检验各处理间的差异,分别对喜旱莲子草克隆片段的端部分株、基部分株和整个克隆片段的生长(总生物量)、克隆繁殖(总匍匐茎长和分株数)和生物量分配(根冠比)进行比较,并运用Duncan检验进行多重比较,若P < 0.05则表示差异显著。所有图示均用Sigmaplot 11.0绘制。
3. 结果
3.1. 端株和基株的克隆繁殖和生长
克隆整合(匍匐茎连接)显著影响喜旱莲子草端株的克隆繁殖(分株数和匍匐茎长)和生长(生物量),不同生态型喜旱莲子草生长和克隆繁殖也差异显著(表1);具体表现为匍匐茎连接显著增加了端株的分株数、匍匐茎长和生物量,并且这种作用在陆生型更为明显(表1;图1)。而对基株部分,匍匐茎连接和生态型都显著影响其克隆繁殖和生长,但两者并无交互作用(表1);匍匐茎连接显著提升了喜旱莲子草基株的所有生长参数,且水生型生长表现明显好于陆生型(表1;图1)。

Figure 1. The growth measures of the apical and basal ramets of Alternanthera philoxeroides, with the stolon connections either severed or kept intact (mean ± SE, n = 8). Different letters indicate significant differences between the treatments (P < 0.05, Duncan’s test)
图1. 不同生态型喜旱莲子草的端株和基株在匍匐茎切断或者连接下的生长参数(均值 ± 标准误,n = 8),不同字母表示处理间差异显著(P < 0.05,Duncan检验)

Table 1. ANOVAs of the effects of stolon connection and different ecotypes on the growth measures of Alternanthera philoxeroides
表1. 匍匐茎连接对不同生态型喜旱莲子草生长参数影响的ANOVA分析表
差异显著的P值用黑体表示;Significant P values are presented in bold。
3.2. 整个克隆片段的生长
在整个克隆片段水平上(端株 + 基株),喜旱莲子草的总生物量积累显著受到匍匐茎连接的影响(F1,28 = 79.21, P < 0.001),并且匍匐茎连接与生态型的交互作用也是显著的(F1,28 = 6.35, P = 0.035)。具体表现为与切断分株相比,匍匐茎连接显著促进了喜旱莲子草克隆片段的生物量积累,并且在陆生型中作用更为显著(图2)。
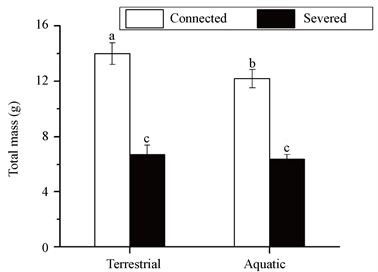
Figure 2. The total biomass of the whole clonal fragment of Alternanthera philoxeroides, with the stolon connections either severed or kept intact (mean ± SE, n = 8). Different letters indicate significant differences between the treatments (P < 0.05, Duncan’s test)
图2. 不同生态型喜旱莲子草的整个克隆片段在匍匐茎切断或者连接下的总生物量(均值 ± 标准误,n = 8),不同字母表示处理间差异显著(P < 0.05,Duncan检验)
3.3. 生物量分配
对于端株部分,仅匍匐茎连接显著影响其根冠比(F1,28 = 19.53, P < 0.001),生态型及其交互作用均不显著;匍匐茎连接显著降低了喜旱莲子草端株的根冠比(图3)。对于基株部分,匍匐茎连接(F1,28 = 78.43, P < 0.001)、生态型(F1,28 = 12.43, P < 0.001)以及两者的交互作用(F1,28 = 7.22, P = 0.012)都显著影响喜旱莲子草的根冠比;具体表现为与匍匐茎切断的分株相比,匍匐茎连接显著增加了基部分株的根冠比,并且在陆生型中作用更为显著(图3)。

Figure 3. The root to shoot ratios of the apical and basal ramets of Alternanthera philoxeroides, with the stolon connections either severed or kept intact (mean ± SE, n = 8). Different letters indicate significant differences between the treatments (P < 0.05, Duncan’s test)
图3. 不同生态型喜旱莲子草的端株和基株在匍匐茎切断或者连接下的根冠比(均值 ± 标准误,n = 8),不同字母表示处理间差异显著(P < 0.05,Duncan检验)
4. 讨论
研究结果很好地支持了我们的假设一,即两种生态型的喜旱莲子草的克隆片段在斑块资源生境中均表现出较为显著的劳动分工现象:以切断分株相比,匍匐茎连接的存在使处于高营养低光照斑块的基部分株分配更多的生物量到根部(根冠比显著则增加)以专化吸收资源较为丰富的地下资源,而处于低营养高光照斑块的端部分株则投入更多的生物量到地上部分(根冠比显著则降低)以专化获取资源较为丰富的光照进而实现克隆扩散 [7]。得益于劳动分工的存在,匍匐茎连接显著促进了两种生态型植株的端部分株、基部分株,以及整个克隆片段的生长(生物量积累)和克隆繁殖(以分株数和匍匐茎长表示)。这很可能是因为植物从资源丰富的环境中获取单位资源所消耗的能量显著小于从资源贫瘠环境中获取等量资源所消耗的能量,因此这种更为经济的资源专化吸收和资源交换可提高整个克隆基株的生产效率,进而使整个基株系统的生物量和适合度也获得增益 [7] [18] [19] [20]。因此,研究结果显示克隆整合及其介导的劳动分工有利于不同生态型喜旱莲子草的生长和克隆繁殖。
此外,我们还发现相比于水生型,陆生型喜旱莲子草克隆片段的基部分株专化地下资源的吸收现象更为明显,即基部分株劳动分工强度更大,而端部分株的专化程度并无显著差异,这一结果部分支持了预测二。自然界中陆生环境的资源分布是普遍异质的,而水生环境由于水流的移动其资源相对较为均一 [17]。陆生型基部植株更强的专化能力表明喜旱莲子草对陆生环境的本地适应性。但有趣的是,相比于水生型,陆生型喜旱莲子草基株更强的专化吸收地下资源能力并没有给予其基株更强的生长表现,而是给予其相连的端部分株更强的克隆繁殖能力和更多的生物量积累。具体体现在匍匐茎连接(克隆整合)更有利于陆生型喜旱莲子草端部分株的克隆繁殖和生长(生长参数水生型增长20%~75%。而陆生型增长39%~120%),从而更强的克隆扩散能力和入侵性。这有可能是因为相对开阔的陆生环境中,相比光照等地上资源,地下资源是相对的限制资源,投入更多的生物量专化获取更多的地下资源,然后通过克隆整合作用与相连分株进行分享能更利于植物的适合度。与之类似,Roiloa等在对外来入侵种莫邪菊的克隆整合作用也研究中发现沙丘和海岸草地两种生态型对异质性资源也有不同程度的响应差异,即异质性更强的沙区生态型的劳动分工能力强于海岸草地生态型,从而给予其更好的生长表现 [7]。
在整个克隆片段水平,克隆整合显著增加了两种生态型的生物量积累,并且陆生型植株获益更大。由于实验设置的负相关的斑块资源是陆生环境,陆生型植株基部更强的专化程度和更显著的生长促进证明了喜旱莲子草的克隆特性如克隆整合和劳动分工能力存在代间可塑性转移(Transgenerational plasticity) [21]。即在陆生更为异质的环境中形成的更强的资源分享能力和劳动分工强度等特性可传至下一代,下一代在陆生环境中会更好的生长表现和适合度。由于喜旱莲子草在国内的基因分化程度极其低 [22] [23],表型可塑性是其应对不同环境的主要适应手段。克隆特性这种代间转移现象有利于喜旱莲子草更好的适应环境。此外,陆生型喜旱莲子草更强的资源分享(克隆整合)能力和劳动分工强度以及由此带来的生长增益可以部分解释为何陆生喜旱莲子草比水生型更加难以进行生物控制:在部分分株遭受天敌取食时,得益于陆生喜旱莲子草由于更强的资源分享和劳动分工能力,未取食分株可以通过克隆整合给予被取食分株更多的支持以缓解昆虫取食压力 [13]。
综上所述,运用实验生态学方法,我们发现,在资源交互的斑块资源条件下,陆生型和水生型喜旱莲子草均体现出明显的劳动分工现象,具体表现为高营养低光照斑块的基部分株分配更多的生物量到根部以专化吸收资源较为丰富的地下资源,处于低营养高光照斑块的端部分株则投入更多的生物量到地上部分以专化获取资源较为丰富的光照进而实现克隆扩散。此外,我们还发现相比于水生型,陆生型喜旱莲子草克隆片段的基部分株专化地下资源的吸收现象更为显著,即基部分株劳动分工强度更大。得益于分株间的劳动分工,匍匐茎连接显著增强了两种生态型的两端分株及其整个克隆片段的生长和克隆繁殖,并且陆生型比水生型喜旱莲子草能更得益于克隆整合,表现为与水生型相比,匍匐茎连接更显著的促进了陆生型端部分株和整个克隆片段的生长和克隆繁殖。值得注意的是,匍匐茎连接显著增加了喜旱莲子草克隆片段的分株数和匍匐茎长(克隆繁殖),而克隆繁殖的增强无疑能增加其无性繁殖体数量,无性繁殖体又可由动物或者人类的介导从成功入侵的种群中扩散至其他区域,进而进一步扩大其入侵范围 [24]。因此,克隆特性(诸如克隆整合和克隆劳动分工)在喜旱莲子草的入侵过程中有着极为重要的作用,特别是对陆生型种群而言。因此,在实际防控喜旱莲子草入侵的过程中,需重视其克隆特性而有针对性的制定措施。
NOTES
*通讯作者。