1. 鸟类迁徙的起源与进化
鸟类迁徙是一个令人着迷的壮观现象。据统计有1855种鸟类(占现存物种的19%)是迁徙的,在它们的繁殖地和越冬地进行有规律的周期性运动,有可预测的时间和目的地 [1]。通常鸟类迁徙的定义是:鸟类在繁殖区和非繁殖区之间有规律的、内源性控制的季节性迁移。因全球气候的变化和人类的活动,预计对鸟类的年度适应性迁徙产生影响 [2] [3] [4],从而出现不适宜的路线和暂时不匹配的繁殖所带来的潜在适应性问题 [5]。因此了解鸟类起源与进化,迁徙时间、研究方法和迁徙影响因素等,对于明确鸟类的生态需求以及保护全球迁徙鸟类资源具有重要的价值,同时也可为预防流行病的传播提供科学依据。
关于鸟类迁徙的起源,通常有三种理论假说:1) “北方家园理论假说”,该假说认为因气候变化,鸟类将它们的非繁殖活动区域从高纬度繁殖地转移出去 [6]。2) “南方家园理论假说”,该假说认为鸟类迁徙是在热带地区进化,但这些热带鸟类迁徙在高纬度地区进行繁殖 [6]。3) Cox GW [7] 和Jahn AE等人 [8] 提出的“近北极–新热带迁徙系统假说”,迁徙的鸟类是温带–热带边缘地区的物种,它们将活动范围扩大到温带繁殖,冬季活动范围向南延伸到热带。但有研究认为,一个物种的祖居地和迁徙进化的起源不必在地理上完全相同 [9]。根据多年来对迁徙鸟类的研究发现,这些假说中某些性状的系统发生起源和进化并不能和地理关联起来,而且这些假说中忽略了鸟类迁徙的多样性,虽然鸟类迁徙大多发生在北半球,但也有鸟类在南半球的温带–热带和热带之间进行迁徙,因而鸟类迁徙的起源还需要科学家对其进行深入研究。
鸟类迁徙进化的关键问题是鸟类通过迁徙,从原来的栖息区域向新的繁殖区域迁移,可以增强繁殖力,提高适应能力。根据对北极燕鸥的研究,发现其是地球上迁徙距离最长的鸟类,每年历经两个夏季,往返于北极北部的繁殖地和南极冰原的栖息地,全部行程长达70,000公里(表1),根据其迁飞模式,提出了一个假设,认为食物分配、风向、换羽习惯等,都是鸟类进化的决定性因素。Helbig AJ [10] 提出:鸟类迁徙是生态影响,而不是系统发育关系影响了一个物种或种群的迁徙行为。
鸟类迁徙在空中没有路标,它们却可以近乎完美的在繁殖地与栖息地往返,因而发现鸟类迁徙是有导航系统的。研究发现,有的鸟类是根据地面上标志性的地貌来导航,如海鸥沿着海岸线进行迁徙;有的鸟类似乎可以感知地球磁场的变化,如信鸽,有实验在其身上安装小磁铁,发现信鸽的方向感紊乱;有的鸟类似乎利用了天文导航的技巧,对特定的天体的光线非常敏感,而这种技巧需要鸟类进行社会学习。大部分迁徙鸟类个体或群体都有固定的迁徙路线和停歇地点,大多鸟类沿着南北方向进行迁徙,也有部分鸟类沿着东西方向迁徙 [11]。全球有八条候鸟迁徙路线,我国地处世界候鸟南北、东西迁徙通道较为关键位置,如图1所示,3、4、5这三条路线与我国鸟类的迁徙有着密切的关系,起中转站的作用。

Table 1. Representative migratory bird information
表1. 具有代表性的迁徙鸟类信息
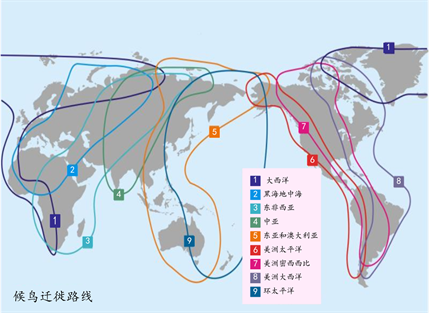
Figure 1. Global migratory bird migration route map. Note: Quoted from the network
图1. 全球候鸟迁徙路线图。注:引自网络
2. 鸟类迁徙研究方法
对于鸟类迁徙的研究通常使用野外观察法、环志法、稳定同位素法 [12]、室内控制实验监测法、无线电追踪、雷达监测法等(表2),但是确定鸟类迁徙使用最多的就是环志法和无线电追踪法,确定鸟类迁徙机制研究通常使用室内控制实验监测法。
Table 2. Advantages and disadvantages of research methods on bird migration
表2. 鸟类迁徙研究方法优缺点
野外观察法:由于对鸟类的认识和了解仍有不足、各地鸟类的名录并不完整、很多鸟类的栖息生境及进食情况记录极少,对于大多鸟类迁徙及季节性繁殖的认识也较为缺乏。鸟类生活在野外,又惧生怕人,因而远距离的野外观察极有必要,通常对鸟类进行野外观察需要做好报告,将观鸟者及同行人的姓名、所记录的种类、观鸟日期、时间、地点、栖息环境(包括大致海拔高度)、天气及光线、距离及观察条件(所使用的助视工具,如单筒望远镜、双筒望眼镜等)、习性记录(飞行、步态、姿势、进食情况及相关种类)、其他情况(观鸟者事前对此种类的熟悉程度、异常天气情况等等)。野外观察法,有助于了解被观测鸟类种群与其栖息环境的关系。
环志法:环志,是对野生鸟类捕捉后测量和搜集基本数据的方法,将捕捉到的鸟类装上标有测量编码的标志物(颈环、翅环、翅旗、脚环),用以标记鸟类,对其进行识别。环志法,最早出现在我国《吕氏春秋》的记载中,而现代的环志研究,始于1899年的丹麦,早期环志采用固定在候鸟足部的刻有编号的金属脚环,现在的环志标志物得到了扩展,不仅使用金属标志物,还使用工程塑料制作编码彩色环。采用最新的环志标志物,可提高环志的回收率并且在不捕捉鸟类的情况下进行识别。最著名的环志鸟类是一只在1980年代初被环志并被媒体以丹麦女王玛格丽特二世名字命名的北极燕鸥(Sterna paradisaea)。稳定同位素法:该技术于上个世纪被引入到生态学研究中的多个领域,稳定同位素携带着生态系统中物质和能量循环过程中的诸多信息,因而成为研究鸟类迁徙的特殊载体和有效手段。稳定同位素,如氢、碳、氮、锶等在空间分布上具有明显的特征差异,赤道附近的氢的稳定同位素氘的比例较高,且这种差异通过食物链传播,积累在鸟类的一些角质化组织上,如羽毛上,积累后能够长期维持其生长时期所在的同位素背景值,而在代谢活跃的组织上,如血液、肌肉等组织仅反映近期的环境特征,因此通过分析迁徙鸟类组织中的稳定同位素含量,来确定鸟类某个时期的分布区域、栖息地特征以及食物类型等特征。稳定同位素法,为了解迁徙鸟类的繁殖地、越冬地及迁徙中途的休息地起到了桥梁作用。
无线电追踪:无线电追踪是近年来随着微电子技术的发展而产生的一种研究鸟类迁徙的方法,它通过安放在鸟类身上的一具无线电信号发射机来精确定位鸟类的位置,从而可以绘制出鸟类迁徙的路线,但是由于技术水平的限制,无线电追踪技术只能应用于鹳、鹤、雕等大型鸟类,无线电跟踪电池寿命短,费用较高,适用于短期个体研究,不适宜群体长期研究。一只名叫Donna的白鹳(Ciconia boyciana)保持着目前无线电追踪世界记录,它于1999年被套上卫星定位发报器,2005年3月5日在法国南部Calvados的塞纳河口触电线身亡,在此期间它为研究者提供了2033天珍贵的科研数据。
雷达监测法:雷达监测法对鸟类迁徙的研究,可以不受天气条件和鸟类飞行的高度影响,可获得飞行时的高度、方向、速度等大量信息。但雷达监测需要多个雷达站进行共同监测,需利用机场及气象站的设备进行合作研究,不仅研究地点的选择受限,贴近地面或海面飞行的鸟类也无法监测到。该方法难以识别被监测到的鸟类的类群信息,只能依靠鸟类个体的大小、飞行速度等信息进行推测。
3. 鸟类迁徙影响因素
3.1. 光周期对鸟类迁徙的影响
通常鸟类的迁徙被认为是鸟类本能,而这种本能不仅仅是由于生理和遗传方面的因素,还有对生存环境长期适应进化的因素。迁徙的鸟类似乎对气候变化很敏感,气候发生了变化,它们开始进行迁飞活动,避免北方冬季的严寒以及南方夏季的酷暑。气候变化对鸟类的食物来源产生了直接的影响,北方秋季天气开始转冷,鸟类的食物开始缺乏,因此鸟类需要迁徙到食物丰美的地区,度过冬天。次年,待北方转暖,繁殖地区食物来源增多,鸟类又返回繁殖地,仅从表面看,鸟类迁徙与食物、气候等生存环境关系极为密切。通过研究得知,鸟类迁徙的主导因素是光周期,光周期是地球上周期变化最稳定的因素,因而也成为许多生物的节律信号,从而对生物的生活史和繁殖周期起到控制作用。鸟类通过光周期确定迁徙时间,这是使用气候和温度因素无法进行解释的。
光周期对鸟类的视觉、神经系统作用,增强了鸟类的内分泌系统活动,如春天日照时间变长,鸟类通过视觉感知光线,作用于下丘脑中枢,抑制睡眠中枢,使得迁徙鸟类处于兴奋状态,增强了垂体活动,促进鸟类生殖腺的活动,为繁殖做准备,影响甲状腺和肾上腺的活动,促使其分泌甲状腺激素和肾上腺激素,增强代谢,存储能量,为迁徙做准备。通过生理准备,机体对外界的刺激更为敏感,当日照长短到了一定的阶段,就激发了鸟类迁飞的本能。秋季日照长度不断缩短,在神经系统和内分泌系统共同作用的情况下,鸟类又迁往栖息地。
3.2. 社会学习对鸟类迁徙的影响
社会学习(Social learning) [13] 通常是指动物个体在群体生活中,通过对其它动物的行为观察或是跟随,获得相关的信息,并改变其行为表现的过程,包括学习和记忆。动物迁徙现象复杂,但其内在机制在鸟类中已有大量的研究,在鸟类迁徙中,有的以个体进行迁徙,如部分猛禽的迁徙,有的则是进行群体迁徙。有的鸟类迁徙是依靠遗传机制,有的鸟类迁徙不仅依靠遗传机制,还需通过社会学习,而且社会学习对迁徙的成功与否至关重要。对于群体迁徙的物种,迁徙信息通常由经验丰富的个体向经验不足的个体传递和交流。
北美体型最大的鸟类:美洲鹤(Grus americana),因其体型高大,叫声高亢而悠远而得名:高鸣鹤,通过对其迁徙的数据分析,发现鸟类迁徙与长期社会学习关系紧密,研究者通过对首次迁徙的高鸣鹤进行人工训练,并使用超轻型飞机保护,发现年龄越大的鸟类对迁徙路径的偏差越小 [14] [15],这是其通过长期社会学习及记忆的结果。
近期有研究通过对长距离迁徙和短距离迁徙的猛禽北极隼(Falco peregrinus)进行种群基因组比较分析,发现了迁徙路线的主要形成原因和长距离迁徙关键基因。该研究通过对北极隼的种群动态推断以及潜在繁殖、越冬地重建发现,在末次冰盛期至全新世的转换过程中,冰川渐退,繁殖地北退和越冬地变迁,可能是北极隼迁徙路线形成的主要历史原因,而且发现环境的差异及适应本土环境在维持迁徙路线中发挥重要作用。该研究还发现,北极隼一个与记忆能力相关的基因ADCY8在长距离迁徙种群中受到了正选择,通过实验证明长、短迁徙种群的主要基因型存在功能差异,因而揭示了长时记忆可能是北极隼长距离迁徙的重要因素之一 [16]。
3.3. 磁场对鸟类迁徙的影响
有研究者对鸟类使用地磁场信息进行导航做了研究,一种是鸟类通过眼睛感知磁场,一种是通过喙感知磁场 [17]。鸟类通过眼睛感知磁场是依据自由基对假说模型,认为蓝光受体隐花色素蛋白(CRY)是磁受体蛋白的候选者,隐花色素在蓝光诱导下可以产生两个电子自旋方向相反或平行的单重态和三重态自由基对,而自由基对具有磁敏感性,在磁场作用下单重态与三重态的比例发生变化从而产生一系列的生理生化过程。隐花色素蛋白CRY是一类进化上保守的黄素蛋白,具有多种生物学功能,在动物体内,CRY起着调节昼夜节律的功能,在鸟类的眼中有四种类型的隐花色素蛋白(CRY):Cry1a、Cry1b、Cry2和Cry4。通过在多个组织中进行研究,发现隐花色素CRY2、时钟蛋白CLOCK,可能在动物的松果体、眼睛中起作用 [18] [19]。基于光受体的生物磁感应自由基对机理是第一个完全吻合动物磁感应特征的理论,Schulten等 [20] 最早提出生物磁感应的内在机理可能是自由基对机理,直到2000年,Ritz等 [21] 才将其发展为完善的生物磁感应自由基对机理假说,并提出隐花色素蛋白是生物体内的磁感应分子。
鸟类通过喙感知磁场是根据磁铁颗粒假说,磁感应细胞能够产生纳米级的呈线性排列的磁小体(磁性颗粒),在运动中感知磁场方向。在动物行为学的研究中发现,磁感应具有许多特征:1) 动物磁感应可以分辨地磁场的磁倾角,但不能分辨磁极方向;2) 动物磁感应具有光波长依赖性,只有在特定波长光照下才能感应地磁场,没有光照和其他波长光照下均不表现磁感应能力;3) 动物磁感应发挥作用具有较窄但并不固定的磁场强度范围 [22]。
4. 小结与展望
鸟类迁徙是高度可变的行为,它将形态、生理、行为、遗传等因素整合。长期以来,鸟类迁徙一直是科学界研究的重点,如野生鸟类禽流感的爆发和蔓延,需要以追溯疫源地为目标对鸟类迁徙进行研究。鸟类迁徙这一行为,距离之远、时间之精确、导航之精准,这些行为特征,不仅对鸟类的保护具有重要的意义,也为其他迁徙物种的研究及仿生学的研究提供了重要的思路。
NOTES
*通讯作者。