1. 引言
光合作用是植物生长发育的基础,是植物光合生产力的重要衡量指标 [1] [2]。不同植物有不同的光适应特性,植物的光合特性除了受植物内在生理因素决定外,也与光、热、水等外界环境因子密切相关 [3] [4] [5]。研究植物光合特性是分析环境因子影响植物生长与发育的重要手段 [6]。
光合作用可以反映植物对环境的适应性,而海拔梯度可以反映出环境因子的综合变化,随着海拔梯度的变化,光照、温度、水分、土壤等环境因子均呈现出规律性的变化 [7],温度、大气压以及二氧化碳浓度等随着海拔的升高而降低,光强、紫外线辐射随海拔的升高而增加 [8]。海拔的变化对环境因子影响显著进而影响植物光合特性。龚荣高等 [9] 研究发现,随着海拔的升高,青脆李最大净光合速率、暗呼吸速率和光补偿点均出现了先增后降的变化。井锐锐等 [10] 研究了4个海拔区域柠檬光合特性,结果为800 m > 1110 m > 420 m > 300 m,即这4个区域中,最适宜种植尤力克柠檬的海拔为800~1110 m。有学者认为高海拔地区的强光环境有利于植物净光合速率的增加 [11] [12],而部分学者持相反观点,认为随着海拔梯度的上升,环境条件会逐渐变得恶劣,高海拔地区的低温、强光、低气压等不利条件限制了植物的生长发育,进而影响植物的光合作用 [13] [14]。这是由于高海拔的低温环境使植物叶片气体交换的能力降低,导致光合能力下降 [15]。由此可见,海拔高度的变化对植物光合特性有显著的影响。因此,研究不同海拔地区植物的光合参数及光响应曲线特征参数,比较其光合特性,对探究不同生态环境下植物的生态适应性具有重要意义。
红松(Pinus koraiensis)在维系东北森林生态系统稳定方面起到重要作用,是我国温带地带性顶极群落小兴安岭阔叶红松林的优势树种 [16] [17]。本研究以黑龙江省丰林国家级自然保护区红松为研究对象,对不同海拔(低海拔、高海拔)样地红松的光合参数及光响应曲线进行测定分析,从光合特性角度探究红松对不同海拔的适应性差异,揭示不同海拔高度对红松光合特性的影响,以期为东北地区红松的经营管理提供科学参考。
2. 材料与方法
2.1. 试验地概况
试验在黑龙江省丰林国家级自然保护区(E128˚58'~129˚15', N48˚02'~48˚12')进行,保护区总面积18,165.4 hm2,其中核心区总面积4165 hm2,缓冲区总面积3812 hm2,试验区总面积10,188.4 hm2,研究区内气候富有大陆性及季风气候相结合的特点,年平均气温为−0.5℃,年平均降水量640.5 mm,空气相对湿度为73%,生长期约100~110天。土壤主要是发育在阔叶红松林下的地带性土壤–暗棕色森林土,通常土层较厚,透水性良好,肥力较高。该区域属长白山植物区系小兴安岭亚区,地带性植被为温带针阔叶混交林,是中国目前保存最完整、最典型的原始红松林保护区。本研究选取不同海拔高度的试验地,分别为低海拔样地和高海拔样地,样地概况见表1。在每个海拔类型内设置3个重复样地,面积为20 m × 30 m,在每个重复样地内随机选取植株健康,长势基本一致的红松3株。
2.2. 研究方法
2.2.1. 光合参数的测定
于2018年6~9月每月中旬选择晴朗无云的上午(8:00~11:30)采用美国lico公司生产的li-6400光合仪进行光合参数的测定,由于无法进行高位活体测量,为保持测量的一致性,所有叶片均进行离体测定,每株测定3束红松,取平均值。测定时,设定叶温为25℃,CO2浓度为400 μmol/mol,湿度为60%,光辐射强度为1500 μmol∙m−2∙s−1。测定净光合速率(Pn, μmol∙m−2∙s−1);气孔导度(Gs, mol∙m−2∙s−1);蒸腾速率(Tr, mmol∙m−2∙s−1);胞间CO2浓度(Ci, μmol/mol−1)。水分利用效率(WUE) = Pn/Tr。
2.2.2. 光响应曲线的测定
使用li-6400光合仪的LED光源,从高到低设置梯度为2000 μmol∙m−2∙s−1、1800 μmol∙m−2∙s−1、1500 μmol∙m−2∙s−1、1200 μmol∙m−2∙s−1、1000 μmol∙m−2∙s−1、800 μmol∙m−2∙s−1、600 μmol∙m−2∙s−1、400 μmol∙m−2∙s−1、200 μmol∙m−2∙s−1、150 μmol∙m−2∙s−1、100 μmol∙m−2∙s−1、50 μmol∙m−2∙s−1、25 μmol∙m−2∙s−1和0 μmol∙m−2∙s−1,设定测量室温度为25℃,CO2浓度为400 μmol/mol,流速为500 μmol/s。记录净光合速率(Pn)。
2.2.3. 数据分析
采用Excel 2019和SPSS 20.0软件对数据分析。本研究光响应曲线使用直角双曲线修正模型计算 [18] [19]。
净光合速率
(1)
(1)式中,Pn为净光合速率;I为光强;α为I = 0时光响应曲线的初始斜率即初始量子效率;β为修正系数;γ = α/Pnmax,Pnmax为最大净光合速率。根据公式可以得出相应的表观量子效率(AQY)、暗呼吸速率(Rd)、最大净光合速率(Pnmax)、光饱和点(LSP)、光补偿点(LCP)等指标 [20] [21] [22]。
3. 结果与分析
3.1. 光合参数差异性比较
3.1.1. 不同海拔红松光合参数差异性比较
不同海拔红松光合参数指标见图1。低海拔和高海拔样地红松的Pn在6~9月份均呈先上升后下降趋势。高海拔红松Pn在6~9月份分别高于低海拔红松4.80%、2.99%、5.33%、4.46%,差异均不显著;在6~9月份低海拔样地Ci浓度分别高于高海拔样地5.63%、8.26%、46.43%、30.20%。8、9月份差异显著;低海拔和高海拔样地红松的Tr在6~9月份均呈现出先上升后下降的趋势,7月份两者差异显著,低海拔样地高于高海拔97.03%,6、8、9月份高海拔样地高于低海拔,差异不显著;低海拔和高海拔样地红松的Gs在6~9月份均呈现先上升后下降的趋势,差异均不显著;低海拔和高海拔样地红松的WUE 7月份差异极显著,高海拔高于低海拔90.85%,6、8、9月份低海拔略高于高海拔,差异不显著。
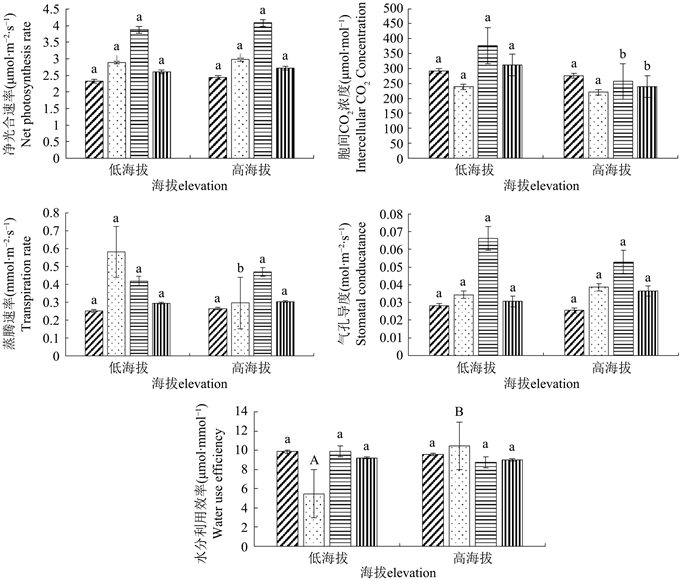
Figure 1. Comparison of photosynthetic parameters of Pinus koraiensis at different elevations. Notes: Uppercase letters in the data label indicate differences at the 0.01 level, and lowercase letters indicate differences at the 0.05 level
图1. 不同海拔红松光合参数比较。注:不同大写字母表示在0.01水平上的差异,不同小写字母表示在0.05水平上的差异
对不同海拔、不同月份红松的光合参数进行方差分析,方差结果见表2,不同海拔红松Ci差异极显著,Pn、Tr、Gs、WUE均无显著差异;不同月份红松Pn、Ci、Tr、Gs均表现出极显著差异,季节变化对红松光合参数的影响显著。

Table 2. Analysis of variance of photosynthetic parameters of Pinus koraiensis at different elevations
表2. 不同海拔红松光合生理指标方差分析
注:**在0.01水平(双侧)上差异显著,*在0.05水平(双侧)上差异显著。
3.1.2. 光合参数相关性分析
低海拔和高海拔红松的光合参数的相互关系见表3。结果表明,Pn与Tr、Gs呈极显著的正相关,Ci与Tr显著负相关,Tr与Gs显著正相关,与WUE呈极显著负相关。
Table 3. Correlation analysis of photosynthetic parameters of Pinus koraiensis
表3. 红松光合参数相关性分析
注:**在0.01水平(双侧)上显著相关,*在0.05水平(双侧)上显著相关。
3.2. 光响应曲线及特征参数差异性比较
3.2.1. 叶子飘模型光强响应拟合效果
光响应曲线反映了植物光合速率随光强改变而变化的规律,对植物光合速率进行客观呈现。采用叶子飘模型对不同海拔红松净光合速率与光照强度之间的关系规律进行拟合,拟合优度大于0.98,结果如图2所示。不同海拔红松6~9月的光响应曲线变化趋势一致,净光合速率随着光强的增加逐渐增加,最后趋于平缓。高海拔红松的净光合速率在6~9月略高于低海拔,差异不显著。
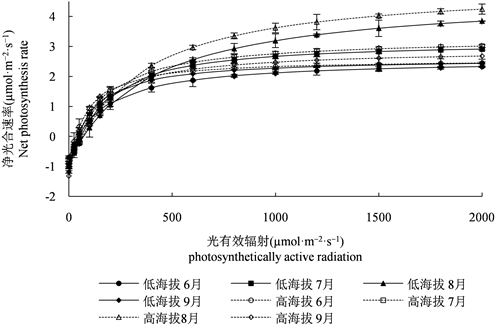
Figure 2. Light response curve fitting effect of Pinus koraiensis
图2. 红松光响应曲线拟合效果
3.2.2. 不同海拔红松光响应曲线特征参数差异性比较
不同海拔红松光响应曲线特征参数差异见表4。低海拔和高海拔样地Pnmax总体上呈先上升后下降的季节动态,在8月份达到最大值。6~9月份高海拔样地红松Pnmax分别高于低海拔样地4.06%、4.14%、6.32%、11.07%,差异显著;低海拔和高海拔样地LSP在6~9月份呈先上升后下降的季节动态,高海拔样地LSP分别高于低海拔样地2.97%、0.35%、4.81%、0.29%,不同海拔间无显著差异;低海拔与高海拔样地LCP在6~9月份呈先上升后下降的季节动态,不同海拔间无显著差异;低海拔与高海拔样地暗呼吸速率总体上呈上升的季节动态,高海拔样地暗呼吸速率在6月份高于低海拔样地2.11%,且差异显著,在8月份高于低海拔样地19.57%,差异极显著;表观量子效率总体上没有明显的季节动态,高海拔样地红松表观量子效率在6月份和8月份高于低海拔样地,且差异显著。
对不同海拔,不同月份红松的光响应特征参数进行方差分析,结果见表5。不同海拔红松间Rd差异显著,Pnmax、LSP、LCP、AQE差异均不显著;不同月份红松的Pnmax、LCP、LSP、AQE、Rd均差异极显著,说明季节变化对红松光合作用影响显著。
Table 4. Characteristic parameters of light response curve of Pinus koraiensis on different elevations
表4. 不同海拔红松光响应曲线特征参数
注:大写字母表示在0.01水平上的差异,小写字母表示在0.05水平上的差异。
Table 5. Variance analysis of characteristic parameters of light response curve of Pinus koraiensis on different elevations
表5. 不同海拔红松光响应曲线特征参数方差分析
注:**在0.01水平(双侧)上差异显著,*在0.05水平(双侧)上差异显著。
3.2.3. 光响应曲线特征参数间的相关性分析
红松光响应曲线特征参数之间的相关性分析见表6。结果表明,Pnmax与LSP及LCP均呈极显著正相关;LCP和LSP呈极显著正相关,与AQE呈极显著负相关;AQE和Rd呈极显著正相关。
Table 6. Correlation analysis between various characteristic parameters of photosynthetic capacity
表6. 光响应曲线特征参数间的相关性分析
注:**在0.01水平(双侧)上显著相关,*在0.05水平(双侧)上显著相关。
4. 结论与讨论
海拔高度的变化对红松光合特性有明显的影响。高海拔红松的光合能力及环境适应能力优于低海拔红松。主要体现在以下几方面:
净光合速率直接反应植物光合作用的强弱,最大净光合速率较高的植物,能积累更多的有机物,植物生长更快 [23]。6~9月份高海拔红松的净光合速率和最大净光合速率均高于低海拔红松,说明高海拔红松具有更强的光合能力,这可能是由于随着海拔的升高,光照强度增强,使叶片能吸收到更多的光能 [11] [12]。
光饱和点高的植物能较好地利用全天且在强光下不易发生光抑制现象。本研究光补偿点和光饱和点随着海拔的高度的增加而升高,说明高海拔红松能更好地利用强光进行光合作用。
植物叶片表观量子效率AQY,反映了植物在弱光阶段对光能的转化利用效率,值越大表明植物叶片利用弱光的能力越强 [24],Rd也与植物的耐弱光性相关,Rd较高说明暗呼吸速率较大,有机物代谢效率较低,不利于有机物的积累。在AQY无明显变化情况下,除6月份外,高海拔红松Rd均低于低海拔红松,表明高海拔红松通过降低暗呼吸作用来间接提高光合性能。
基金项目
国家自然基金面上项目(41275154);黑龙江省森林工业总局科技计划(sgzjQ2015006; sgzjY2018001);黑龙江省财政自拟课题(2015-02SCZ);黑龙江省自然科学基金资助项目(LH2020C104)。
NOTES
*第一作者。
#通讯作者。