摘要: 为研究氮素营养对黄檗幼苗生长和光合生理的影响,以当年生水培黄檗幼苗为实验材料,在0、2、8、16 mmol•L
−1四种不同氮浓度条件下,探讨不同氮素营养对黄檗幼苗生长、光合作用和叶绿素荧光等的影响。结果表明,随着氮浓度的提高,黄檗幼苗的鲜重、干重、基径和侧根长度、净光合速率(P
n)、表观量子效率(AQY)、光饱和点(LSP)、气孔导度(G
s)和蒸腾速率(T
r)均呈先上升后下降的趋势,且均在8 mmol•L
−1处理时达到最大值,而光补偿点(LCP)则呈持续下降趋势。最大荧光(F
m)、PSII最大光化学效率(F
v/F
m)、PSII反应中心光捕获效率(F
v'/F
m')、快速叶绿素荧光诱导曲线(OJIP曲线)均以氮浓度为8 mmol•L
−1时最高。不同施氮素水平(N2、N8和N6)下,黄檗幼苗叶片的光化学淬灭系数(qP)均明显高于不施氮素水平(N0) (P < 0.05),并随着氮浓度的增加,qP呈降低趋势;而非光化学淬灭系数(NPQ)则整体呈增加趋势。综合上述研究内容,氮素浓度在8 mmol•L−1时最有利于黄檗幼苗的生长且可以充分发挥其光合能力,这对黄檗苗木资源培育营养管理与调控提供了指导意义。
Abstract:
In order to study the effects of nitrogen nutrition on the growth and photosynthetic physiology of Phellodendron amurense seedling, the hydroponic seedling of P. amurense was used as experimental material, the effects of different nitrogen nutrition on the growth, photosynthesis and chlorophyll fluorescence of P. amurense seedlings were investigated under the conditions of 0, 2, 8, 16 mmol•L−1. The results showed that with the increase of nitrogen concentration, the fresh weight, dry weight, base diameter and lateral root length, net photosynthetic rate (Pn), apparent quantum efficiency (AQY), light saturation point (LSP), stomatal conductance (Gs) and transpiration rate (Tr) of P. amurense seedlings were firstly increased and then decreased, and all reached the maximum value when the nitrogen concentration was 8 mmol•L−1. While the light compensation point (LCP) showed a trend of continuous decreasing with the increase of nitrogen concentration. Maximum fluorescence (Fm), maximum photochemical efficiency of PSII (Fv/Fm), photo capture efficiency of PSII reaction center (Fv'/Fm') and rapid chlorophyll fluorescence induction curve (OJIP curve) were the highest when nitrogen concentration was 8 mmol•L−1. Under different nitrogen application levels (N2, N8 and N6), the photochemical quenching coefficient (qP) of leaves of P. amurense seedlings was significantly higher than that without nitrogen application (P < 0.05), and qP showed a decreasing trend with the increase of nitrogen concentration (N0). However, the non-photochemical quenching coefficient (NPQ) showed an overall increasing trend. In conclusion, the nitrogen concentration of 8 mmol•L−1 was found to be the most favorable for the growth of P. amurense seedlings and to give full play to their photosynthetic capacity, which provided guidance for the nutrient management and regulation of P. amurense resources cultivation.
1. 引言
氮素是植物生长过程中的必需营养元素之一,也是构成植物体内许多重要有机化合物的主要成分。氮素在植物营养生理中也起着非常重要的作用,供氮水平可通过影响氮代谢关键酶活性而影响植物的生长、发育以及对环境的适应。除了少数的植物可以利用固氮微生物吸收空气中的氮外,大多数植物都是通过根系吸收土壤中的无机态氮。氮素供应水平对植物体内的硝酸还原酶和谷氨酰胺合成酶活性有不同程度的影响;也在一定程度上影响可溶性蛋白含量,而可溶性蛋白大部分为参与各种代谢的酶类,是植物体内贮藏氮的主要形式,其中绝大多数是卡尔文循环中最关键的酶,在植物的光合作用、生长发育及植物生物量的建成中起着重要作用 [1] [2]。光合作用是植物生长的重要能量来源和物质基础,氮素浓度能够显著影响植物的生长和光合作用,主要影响以下特征:叶面积、叶片比面重、叶片氮素含量和Rubisco酶活性等。另外,不同氮素浓度还可以通过改变气孔导度和光呼吸速率等因素来调控光合速率。
黄檗(Phellodendron amurense)是芸香科(Rutaceae)黄柏属(Phellodendron)高大乔木,又称黄波萝,主要分布于我国东北地区大兴安岭南部、小兴安岭、长白山、完达山、千山及华北地区燕山山脉以北的山区,在俄罗斯远东和萨哈林南部、朝鲜、日本也有少量的分布 [3]。黄檗是我国东北“三大硬阔”之一,也是我国药典收录的传统大宗药材品种——关黄柏的唯一来源植物。因关黄柏具有清热燥湿,泻火除蒸,解毒疗疮之功效 [4],市场对关黄柏的需求量逐年增加,作为关黄柏的唯一来源植物的黄檗目前仍只能依赖于天然资源,但现有野生资源日益减少,急需大规模人工培育优良资源,以保护现有天然黄檗资源 [5]。有研究表明,适量施入N肥对黄檗苗木生长有促进作用 [6] [7],本文在前人基础上主要研究了不同氮素水平对黄檗幼苗生长、光合生理特性和叶绿素荧光的影响,确定黄檗生长所需的最佳氮素条件,以便进一步对黄檗资源培育和主要药用成分含量的氮肥管理与调控提供基础数据。
2. 材料与方法
2.1. 实验材料
试验在东北林业大学植物光照培养室中进行,温度设定为18℃~25℃,每日光照时间为早7点晚5点。将打破休眠的黄檗种子消毒后播种于托盘中,在黄檗幼苗生长到两片真叶时,挑选长势均匀一致的幼苗移栽至不透光的培养盆中进行水培,用具有孔隙的塑料板将幼苗固定。幼苗先以自来水培养2天,再以50%浓度的完全Hoagland营养液培养一周,然后再用完全营养液培养一周,使幼苗生长健壮,去除弱苗、保留长势一致的壮苗用于进一步的研究。
完全营养液按文献 [8] 的方法配制,略加修改:NH4NO3 (4.0 mmol/L)、KH2PO4 (1.0 mmol/L)、KC1 (1.0 mmol/L)、CaCl2·6H2O (1.0 mmol/L)、MgSO4·7H2O (0.6 mmol/L)、FeCl3·6H2O (0.02 mmol/L)、MnCl2·4H2O (6.0 μmol/L)、H3BO3 (0.016 mmol/L)、ZnCl2 (0.3 μmol/L)、CuCl2·2H2O (0.3 μmol/L)、Na2MoO4·2H2O (0.3 μmol/L)。
2.2. 实验处理方法
在完全营养液配方的基础上,通过改变NH4NO3的浓度配制出4个氮浓度水平:0、2、8和16 mmol/L (分别标记为N0、N2、N8和N16)。每个处理设3次重复,营养液3天更换1次,全天用泵通气,以防止黄檗苗根部发生霉变和腐烂。每天用0.1 mmol/L的KOH和HCl将营养液pH值调至6.0左右,并相应地补充损失的水分;在不同氮素浓度处理3个月后测定不同处理幼苗的各项生理参数。
2.3. 光响应曲线的测定
利用便携式光合仪(Li-6400,美国LI-COR公司),将仪器自带光源分别设定光强(PFD)为1800、1500、1200、1000、800、500、200、100、50和0 μmol·m−2·s−1,按照光强由高到低的顺序分别测定不同氮浓度处理下黄檗幼苗叶片的净光合速率(Pn)、气孔导度(Gs)和蒸腾速率(Tr),并分别绘制Pn-PFD、Gs-PFD和Tr-PFD响应曲线,用直角双曲线模型求得Pn-PFD响应曲线表观量子效率(AQY)、暗呼吸速率(Rd)、光补偿点(LCP)、光饱和点(LSP)、光饱和时的净光合速率(Pmax)。
2.4. 叶绿素荧光参数的测定
采用便携式脉冲调制荧光仪(FMS-2,英国Hansatch公司),利用暗适应夹对不同处理下黄檗幼苗顶端第2片完全展开叶片进行0.5 h的暗适应,参照Hu等 [9] 方法测定初始荧光(Fo)、最大荧光(Fm)和可变荧光(Fv),以及光适应下的最大荧光(Fm')、光适应下的最小荧光(Fo'),计算PSII最大光化学效率(Fv/Fm)、PSII反应中心光能捕获效率(Fv′/Fm′)、光化学淬灭系数(qP)和非光化学淬灭(NPQ)等。每处理3次重复。
2.5. 快速叶绿素荧光动力学曲线(OJIP)的测定
选择不同氮水平处理下黄檗幼苗完全展开叶片,进行30 min暗适应后利用Mini调制式掌上叶绿素荧光仪(Fluorpen FP 100 max,捷克)测定各暗适应后叶片的OJIP曲线。OJIP曲线由3000 μmol·m−2·s−1的脉冲红光诱导,相对荧光强度的记录从10 μs开始至1 s结束,利用5次重复的平均值绘制OJIP曲线。其中OJIP曲线上O、J、I和P点分别为0、2、30和1000 ms对应的时刻,L和K点分别表示0.15和0.3 ms对应的时刻。
2.6. 数据处理
每个实验重复处理3次。采用Excel 2010、Sigmaplot 12.5等软件进行绘图、利用SPSS 18.0软件进行方差分析、LSD法进行差异显著性分析,其中,显著性水平P < 0.05,以小写字母表示其显著差异性;显著性水平P < 0.01,以大写字母表示其极显著差异性。
3. 结果与分析
3.1. 不同氮浓度对黄檗幼苗植株生长的影响
 Note: Lowercase letter and capital letter mean the differences are significant (P < 0.05) and (P < 0.01) respectively, same below.注:同一参数中不同小写字母表示差异显著(P < 0.05),不同大写字母表示差异极显著(P < 0.01),下同。
Note: Lowercase letter and capital letter mean the differences are significant (P < 0.05) and (P < 0.01) respectively, same below.注:同一参数中不同小写字母表示差异显著(P < 0.05),不同大写字母表示差异极显著(P < 0.01),下同。
Figure 1. Effects of different nitrogen concentration on growth of P. amurense seedlings
图1. 不同氮浓度对黄檗幼苗植株生长的影响
由图1可以看出,随着营养液中氮浓度的增加,黄檗幼苗的地下部、地上部鲜重和总鲜重以及地下部、地上部干重和总干重均呈先增加后降低的趋势,并且均在氮浓度为8 mmol·L−1时达最大值,氮浓度过高(16 mmol·L−1)时,黄檗幼苗的各部位鲜重和干重均稍低于氮浓度为2 mmol·L−1时,但不同氮浓度下黄檗幼苗的生长均明显高于无氮处理。不同氮浓度处理下黄檗幼苗的直径和根长与鲜重和干重的变化趋势相似,氮浓度为8 mmol·L−1时的直径分别高于氮浓度为0、2和16 mmol·L−1的19.12% (P < 0.05)、10.81% (P > 0.05)和8.86% (P > 0.05),而根长分别高于0、2和16 mmol·L−1的17.73% (P > 0.05)、6.82% (P > 0.05)和28.52% (P > 0.05)。
3.2. 不同氮浓度对黄檗幼苗叶片光合特性的影响
3.2.1. 不同氮浓度对黄檗幼苗叶片净光合速率光响应曲线的影响
由图2可以看出,不同氮浓度下黄檗幼苗叶片的净光合速率Pn-PFD曲线明显不同,其中光强为0时,不同处理下黄檗幼苗叶片的Pn差异较小,即暗呼吸速率Rd差异较小,但不同浓度处理下以氮浓度为16 mmol·L−1时Rd最高,而氮浓度为0 mmol·L−1最低。随着光强的增加,氮浓度为0 mmol·L−1时黄檗幼苗叶片的Pn增加幅度较小。随着光强的增加,氮浓度为8 mmol·L−1时黄檗幼苗叶片的Pn增加幅度最大,氮浓度为2 mmol·L−1次之。
根据直角双曲线模型求得Pn-PFD响应曲线的各光合参数可以看出(表1),表观量子效率AQY表现为N8 > N2 > N16 > N0,光饱和点LSP表现为相同的变化趋势,但黄檗幼苗叶片的光补偿点LCP则随着营养液中氮浓度的增加呈降低趋势。不同氮浓度处理下黄檗幼苗叶片的最大净光合速率Pmax在氮浓度为8 mmol·L−1时最高,氮浓度2 mmol·L−1次之,明显高于氮浓度为16和0 mmol·L−1时,并且而营养液中氮浓度为0 mmol·L−1时黄檗幼苗叶片的Pmax最低。

Figure 2. Effects of different nitrogen concentration on net photosynthetic rate to illumination intensity in leaves of P. amurense seedlings
图2. 不同氮浓度对黄檗幼苗叶片净光合速率光响应曲线的影响

Table 1. Photosynthetic parameters in leaves of P. amurense seedlings under different nitrogen concentration
表1. 不同氮浓度对黄檗幼苗叶片的光合参数
3.2.2. 不同氮浓度对黄檗幼苗叶片气孔导度(Gs)和蒸腾速率(Tr)光响应曲线的影响
由图3可以看出,不同氮浓度下黄檗幼苗叶片的Gs和Tr的变化趋势相似,随着光强的增加,不同处理下黄檗幼苗叶片的Gs和Tr均有小幅度的增加,但随光强增加,氮浓度为2和8 mmol·L−1时黄檗幼苗叶片的Gs和Tr增加的幅度明显大于氮浓度为0和16 mmol·L−1时。另外,在不同光强下,不同氮浓度处理下黄檗幼苗叶片的Gs和Tr均表现为N8 > N2 > N0 > N16,其中N8和N2处理较为接近,并且明显大于N0和N16处理。
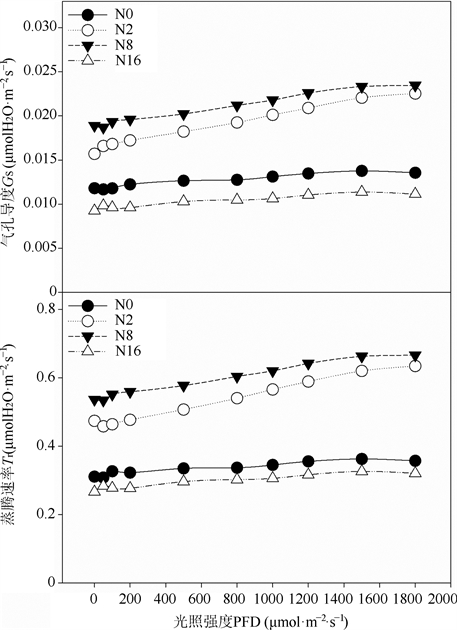
Figure 3. Effects of different nitrogen concentration on stomatal conductance and transpiration rate to illumination intensity, in leaves of P.amurense seedlings
图3. 不同氮浓度对黄檗幼苗叶片气孔导度和蒸腾速率光响应曲线的影响
3.3. 不同氮浓度对黄檗幼苗叶片叶绿素荧光特性的影响
3.3.1. 不同氮浓度对黄檗幼苗叶片叶绿素荧光参数的影响
由图4可以看出,在营养液中氮浓度为0 mmol·L−1时黄檗幼苗叶片的最大荧光(Fm)、PSII最大光化学效率(Fv/Fm)和PSII反应中心光能捕获效率(Fv′/Fm′)均明显低于不同氮素浓度处理,氮浓度为2、8和16 mmol·L−1时黄檗幼苗叶片的Fm分别高于氮浓度为0 mmol·L−1时70.29%、79.82%和75.50%,均达极显著差异水平(P < 0.01),Fv/Fm分别高于氮浓度为0 mmol·L−1时14.86% (P < 0.05)、20.76% (P < 0.01)和20.24% (P < 0.01),Fv′/Fm′分别高于氮浓度为0 mmol·L−1时34.42% (P < 0.01)、37.76% (P < 0.01)和24.86% (P < 0.01),其中,氮浓度为8 mmol·L−1时黄檗幼苗叶片Fm、Fv/Fm和Fv′/Fm′最高。

Figure 4. Effects of different nitrogen concentration on chlorophy II fluorescence characteristics in leaves of P. amurense seedlings
图4. 不同氮浓度对黄檗幼苗叶片叶绿素荧光特性的影响
3.3.2. 不同氮浓度对黄檗幼苗叶片非光化学淬灭(NPQ)和光化学淬灭系数(qP)的影响
由图5可以看出,当氮浓度2和8 mmol·L−1时,黄檗幼苗叶片的NPQ与氮浓度为0 mmol·L−1时的差异较小,但当氮浓度增加到16 mmol·L−1时却分别较0、2和8 mmol·L−1的氮浓度下黄檗幼苗叶片的NPQ增加了76.77% (P < 0.05)、93.39% (P < 0.05)和54.85% (P > 0.05),与NPQ不同,在2、8和16 mmol·L−1的氮浓度下黄檗幼苗叶片qP均明显高于氮浓度为0 mmol·L−1时,但随着营养液中氮浓度的增加,黄檗幼苗叶片的qP呈明显的降低趋势。
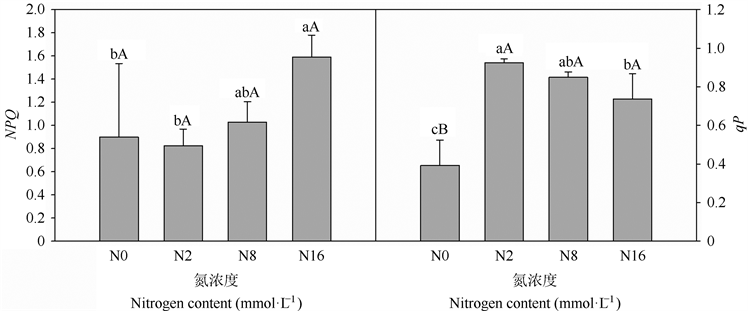
Figure 5. Effects of different nitrogen concentration on qP and NPQ in leaves of P. amurense seedlings
图5. 不同氮浓度对黄檗幼苗叶片光化学淬灭系数(qP)和非光化学淬灭(NPQ)的影响
3.3.3. 不同氮浓度对黄檗幼苗叶片OJIP曲线的影响
由图6可以看出,不同氮浓度处理下黄檗幼苗叶片的OJIP曲线各点的相对荧光强度明显不同,其中,整体来看各点的相对荧光强度以氮浓度为8 mmol·L−1时最高,氮浓度为2和16 mmol·L−1处理下黄檗幼苗叶片各点的相对荧光强度均明显降低,而在氮浓度为0 mmol·L−1时,O点到J点(即0 ms~2 ms)的相对荧光强度稍高于其它3个供氮水平,而超过2 ms时,黄檗幼苗叶片各点的相对荧光强度明显低于氮浓度为8 mmol·L−1时,却高于氮浓度为2和16 mmol·L−1时。
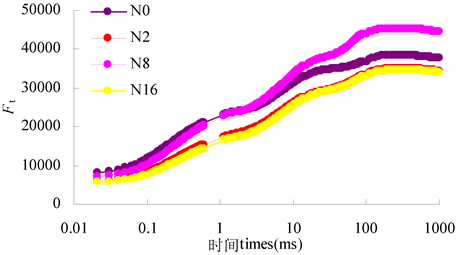
Figure 6. Effects of different nitrogen concentration on OJIP curve in leaves of P. amurense seedlings
图6. 不同氮浓度对黄檗幼苗叶片OJIP曲线的影响
4. 讨论
氮是植物体内最为重要的营养元素之一,在植物的生长发育和代谢过程中占有重要的地位 [10]。合理的氮素调控是保证植物正常生长发育的重要措施之一 [11] [12]。本研究结果表明,不同氮水平下黄檗幼苗植株的生物量积累明显不同,其中,在无氮素供应时,黄檗幼苗的地上部、地下部鲜重和干重均最低,并且其植株的直径和根系长度也明显低于供氮处理。但是,随着营养液中氮浓度的增加,黄檗幼苗植株的地上部、地下部生物量以及鲜重均呈现先增加后降低的趋势,并且不同处理下均以氮浓度为8 mmol·L−1时达最大值,即黄檗幼苗水培条件下较适宜的供氮水平约为8 mmol·L−1,当氮素浓度过高或过低时均会抑制黄檗幼苗的生长。
光合作用是植物获取物质和能量的基础,氮作为植物叶绿素的重要组成成分,其供应能力直接影响着植物对光能的吸收和利用,因此,氮除了直接影响植物的生长及氮代谢过程外还会间接影响植物的光合作用 [13]。碳氮代谢是植物生命活动中最重要的两大代谢过程,二者常偶联在一起发挥作用 [14]。本研究中,不同氮素水平下黄檗幼苗叶片的净光合速率Pn在不同光强下明显不同,其中不同氮水平下以氮浓度为8 mmol·L−1时黄檗幼苗叶片的Pn最高,而以氮浓度为0 mmol·L−1最低,说明无氮营养供应时明显抑制了黄檗幼苗叶片的碳同化能力和光能利用能力,其原因可能与无氮时黄檗幼苗叶片的叶绿素合成受到抑制,导致其对光能的捕获能力明显降低,或氮素供应不足导致其光合作用相关酶的合成或活性受到抑制有关,但这方面的验证有待更进一步的研究。另外,氮素供应过高时,即氮浓度为16 mmol·L−1时黄檗幼苗叶片的光合能力也受到了限制,这与魏丽娜等人在水曲柳上的研究结果相似 [15]。光合曲线拟合出的参数表观量子效率(AQY)是植物对光能利用效率的重要指标,反映了植物捕获光量子用于光合作用的能力 [16],光饱合时的最大光化学效率(Pmax)体现了植物潜在的光合能力 [17],光补偿点和光饱合点(LCP和LSP)反映了植物对光能的利用能力。本研究中,根据直角双曲线模型求得Pn-PFD响应曲线的各光合参数可以看出,黄檗幼苗叶片的AQY和LSP均表现为N8 > N2 > N16 > N0,而黄檗幼苗叶片的光补偿点LCP则随着营养液中氮浓度的增加呈降低趋势,即在无氮供应时,黄檗幼苗叶片对光能的捕获和利用能力明显降低,特别是对弱光的捕获能力和对强光的利用能力下降是导致其光合碳同化能力降低的主要原因之一。不同氮浓度下黄檗幼苗叶片的气孔导度(Gs)和蒸腾速率(Tr)的变化趋势相似,随着光强的增加,不同处理下黄檗幼苗叶片的Gs和Tr均有小幅度的增加,但随光强增加,氮浓度为8和2 mmol·L−1时黄檗幼苗叶片的Gs和Tr增加的幅度明显大于氮浓度为0和16 mmol·L−1时。另外,在不同光强下,不同氮浓度处理下黄檗幼苗叶片的Gs和Tr均表现为N8 > N2 > N0 > N16,其中N8和N2处理较为接近,并且明显大于N0和N16处理。说明在合理的氮素供应下可以保证黄檗幼苗叶片的气孔开度和正常的蒸腾速率,以保证碳同化原料CO2的正常供应和水分代谢过程。
叶绿素荧光技术作为研究植物光合作用的重要手段之一,在反应植物光合作用内在机理,特别是PSII光化学活性和光能利用方面发挥着重要的作用 [18] [19]。本研究中,不同氮素水平下黄檗幼苗叶片的最大荧光(Fm)、PSII最大光化学效率(Fv/Fm)和PSII反应中心光能捕获效率(Fv′/Fm′)均明显高于无氮素供应时,即氮素的供应会明显影响黄檗幼苗叶片的PSII光化学效率,但是不同供氮水平下,黄檗幼苗叶片的Fm、Fv/Fm和Fv′/Fm′均以营养液氮浓度为8 mmol·L−1时最高,即此时黄檗幼苗叶片的PSII光化学效率最高,氮素供应过高或过低时均会抑制其PSII光化学效率,这与光合碳同化的结果相符合,这也是导致不同处理之间黄檗幼苗叶片光合碳同化之间差异的内在原因。光化学淬灭系数(qP)在一定程度上反应了光系统II反应中心的开放程度 [20],而Li等人的研究发现,非光化学淬灭(NPQ)则与依赖叶黄素循环的热耗散显著正相关,即NPQ主要为依赖叶黄素循环的能量耗散部分 [21]。本研究中,不同氮素水平下黄檗幼苗叶片的qP均明显高于不施氮水平,即氮素可以明显促进黄檗幼苗叶片的PSII反应中心光开放程度,但随着氮浓度的增加,黄檗幼苗叶片的qP呈降低趋势,而NPQ则整体呈增加趋势,说明较高氮浓度下,黄檗幼苗叶片的过剩光能主要通过依赖于叶黄素循环的能量耗散方式进行耗散,以减轻PSII反应中心的压力,但在无氮供应时,由于NPQ水平较低,过剩光能可能不能进行有效耗散,从而导致其PSII反应中心光开放程度和光化学活性较低。
不同氮水平下均明显改变了黄檗叶片的快速叶绿素荧光诱导动力学曲线(OJIP)曲线形态,通过对不同处理下黄檗叶片的OJIP曲线标准化分析可以看出,不同氮素水平下黄檗叶片的J点的相对可变荧光呈明显不同,其中,氮浓度为0 mmol·L−1时黄檗幼苗叶片J点和I点的相对可变荧光最高,但J点相对可变荧光的变化幅度明显大于I点。J点的相对可变荧光可以反映电子传递体QA的还原量,即QA−的积累量,J点的相对可变荧光的增加被认为是光合电子传递链中QA (光系统II电子传递链初级电子受体)到QB (光系统II电子传递链次级电子受体)的电子传递受阻,还原态QA积累量逐渐增多,而I点荧光变化则反映了PQ库的异质性 [22] [23],即在无氮素供应时,黄檗幼苗叶片PSII电子传递链电子受体侧的电子传递受阻,表现为PSII受体侧接受电子的能力降低,但其主要抑制作用位点主要发生在QA向QB的电子传递过程,即QB接受电子能力的降低,而受PQ库的影响相对较小。另外,在氮浓度为8 mmol·L−1时黄檗幼苗叶片J点和I点的相对可变荧光最低,而氮浓度为2和16 mmol·L−1时的变化较小,即此时对黄檗幼苗PSII受体侧电子传递速率的影响较小,而光合PSII活性的降低和功能的抑制可能与其它因素有关。
5. 结论
黄檗幼苗的重量、基径和侧根长度、净光合速率(Pn)、表观量子效率(AQY)、光饱和点(LSP)、气孔导度(Gs)和蒸腾速率(Tr)等光合指标,最大荧光(Fm)、PSII最大光化学效率(Fv/Fm)、PSII反应中心光捕获效率(Fv′/Fm′)、快速叶绿素荧光诱导曲线(OJIP曲线)等叶绿素荧光指标均在氮深度为8 mmol·L−1处理时达到最大值,表明氮素浓度在8 mmol·L−1时最有利于黄檗幼苗的生长和壮苗培育,这种对生长促进作用主要是通过提升黄檗幼苗的光合和叶绿素荧光能力、体内碳氮代谢来实现的。研究结果有助于黄檗的资源培育。
基金项目
感谢黑龙江省应用技术研究与开发计划项目(GA19B201)和国家自然科学基金项目(31370630)的资助。
NOTES
*通讯作者。