胚胎干细胞的分化和多能干细胞巢模型——动物经络系统中胚胎干细胞的分化
Embryonic Stem Cell Differentiation and Pluripotent Stem Cell Niche Model—Differentiation of Embryonic Stem Cells in Animal Meridian System
摘要:
经络系统是定向干细胞、成体干细胞、多能干细胞和胚胎干细胞等活动交流、协同进化的巨系统,主要表现为干细胞巢的出现以及不同种类干细胞巢的有序分布。本文首先论述了满足生物重演律要求的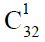 (造血干细胞是中的成员)、
(造血干细胞是中的成员)、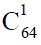 (髓系祖细胞和淋巴系祖细胞是中的成员)和
(髓系祖细胞和淋巴系祖细胞是中的成员)和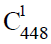 多能干细胞巢模型,其次阐述了动植物进化树之间普遍存在的对应关系以及动植物细胞组织的分化,指出维管植物中的两条细胞分化链,决定了孢子体与配子体的世代交替以及生殖器官(被子植物的花)的形成与演化,因为维管植物的孢子体与配子体先后参与构建一条长度始终不变的生殖细胞分化链,所以孢子体世代越发达,配子体世代就越简化。同理,在脊椎动物的经络系统中,同样以全能细胞ES胚胎干细胞(约定用
多能干细胞巢模型,其次阐述了动植物进化树之间普遍存在的对应关系以及动植物细胞组织的分化,指出维管植物中的两条细胞分化链,决定了孢子体与配子体的世代交替以及生殖器官(被子植物的花)的形成与演化,因为维管植物的孢子体与配子体先后参与构建一条长度始终不变的生殖细胞分化链,所以孢子体世代越发达,配子体世代就越简化。同理,在脊椎动物的经络系统中,同样以全能细胞ES胚胎干细胞(约定用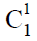 表示)和EG胚胎干细胞(约定用
表示)和EG胚胎干细胞(约定用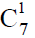 表示)为起点的两条干细胞分化链
表示)为起点的两条干细胞分化链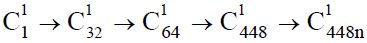 和
和 ,决定了、和多能干细胞巢的形成和经络系统内的板块演化。
,决定了、和多能干细胞巢的形成和经络系统内的板块演化。
Abstract:
The meridian system is a giant system of active communication and co-evolution of human committed stem cells, adult stem cells, pluripotent stem cells, and embryonic stem cells. It is mainly manifested by the appearance of stem cell niches and the orderly distribution of diverse stem cell niches. This paper first describes (such as hemopoietic stem cells), (such as myeloid progenitors and lymphoid progenitors), and pluripotent stem cell niche models that meet the requirement of biological recapitulation laws, then explains the evolutionary relationships and tissue differentiation of animals and plants in an evolutionary tree, and finally points out two cell differentiation chains of vascular plants. The two cell differentiation chains of vascular plants determine the generational alternation of sporozoites and gametophytes and the formation and evolution of reproductive organs (flowers of angiosperms). Because the sporophytes and gametophytes of vascular plants are involved in the construction of a germline differentiation chain with a constant length, so the more advanced the sporophyte generation, the simpler the gametophyte generation will be. Similarly, in the vertebrate meridian system, the two stem cell differentiation chains, and , which are based on pluripotent ES embryonic stem cells (represented by) and EG embryonic stem cells, (represented by) respectively, determines the formation of ,and pluripotent stem cell niches and the plate evolution in the meridian system.
参考文献
|
[1]
|
Zhang, J.X. (2016) Evolutionary Mechanisms for Stem Cell Differentiation and Stem Cell Niche Distribution. Chinese Journal of Tissue Engineering Research, 20, 7571-7578. [Google Scholar] [CrossRef]
|
|
[2]
|
Zhang, J.X. (2018) Main Cellular Components and Their Combination Models of Stem Cell Niches. Advances in Clinical Medicine, 8, 694-701. [Google Scholar] [CrossRef]
|
|
[3]
|
Levit, G.S., Hossfeld, U. and Olsson, L. (2015) Alexei Sewertzoff and AdolfNaef: Revising Haeckel’s Biogenetic Law. History & Philosophy of the Life Sciences, 36, 357-370. [Google Scholar] [CrossRef] [PubMed]
|
|
[4]
|
Wedgwood, K.C.A. and Satin, L.S. (2018) Six Degrees of Depolarization: Comment on “Network Science of Biological Systems at Different Scales: A Review” by Marko Gosak et al. Physics of Life Reviews, 24, 136-139. [Google Scholar] [CrossRef] [PubMed]
|
|
[5]
|
Lópezotín, C., Blasco, M.A., Partridge, L., et al. (2013) The Hallmarks of Aging. Cell, 153, 1194-1217. [Google Scholar] [CrossRef] [PubMed]
|
|
[6]
|
Coito, J.L., Silva, H., Ramos, M.J.N., et al. (2018) Vitis Flower Sex Specification Acts Downstream and Independently of the ABCDE Model Genes. Frontiers in Plant Science, 9, 1029. [Google Scholar] [CrossRef] [PubMed]
|
|
[7]
|
Yang, Y., Fu, D.Z. and Wang, Q. (2004) Origin of Flowers: Hypotheses and Evidence. Acta Botanica Boreali-Occidentalia Sinica, 24, 2366-2380. [Google Scholar] [CrossRef]
|