1. 引言
普通野生稻(Oryza rufipogon Griff.)是栽培稻(Oryza sativa L.)的近缘种,遗传多态性较栽培稻丰富 [1] [2] [3]。其中蕴含不少抗病虫害、耐非生物逆境、具高产数量性状位点、米质优良、雄性不育的优、特异资源 [4] - [11],是改良栽培稻以提高其产量、品质、抗性与适应性的重要遗传资源。有关野生稻光合作用的研究报道不多 [12] - [18],而有关低温对野生稻光合作用影响的报道更少,海南野生稻是普通野生稻的一个种群,有关低温对其光合作用的影响未见报道。本研究比较分析了海南野生稻与典型栽培稻在低温胁迫后的净光合速率(光合强度)、气孔导度、表观量子效率、羧化效率、光系统II的最大光化学量子效率与光合色素含量,旨在探讨海南野生稻光合作用对低温的忍耐能力,发掘其适应低温的优良光合生理指标性状,为其开发利用提供理论依据并获取有关野生稻光合作用的新知识,具有重要意义。
2. 材料与方法
2.1. 材料
海南野生稻(Oryza rufipogon Griff.)、日本晴(Oryza sativa L. ssp. japonica)与9311 (Oryza sativa L. ssp. indica)。
2.2. 方法
2.2.1. 材料栽培与管理
供试材料用种茎于2010年9月下旬移栽,种茎高20 cm,每桶栽2个种茎,每个种茎带2个分蘖,每材料栽3盆。盆中装有湿润水稻土(水稻土烘干重为
3.5 Kg
),自然条件下生长发育,常规水肥管理与病虫防治,至分蘖数足够时(约3周)进行各项光合指标参数的测定。
2.2.2. 低温胁迫
利用长沙晚秋的自然低温对材料进行低温胁迫。低温胁迫日期为
2010 年 10 月 26 日
至2010年10月31日,低温胁迫期间平均日均温为12.57℃,平均日最低温为9.37℃,平均日最高温为16.60℃,低温胁迫后,在自然条件下恢复生长5天(
2010 年11 月1 日~
2010 年11 月5 日
,5天平均日均温为15.50℃,平均日最低温为11.62℃,平均日最高温为21.16℃)后,用于低温胁迫后光合作用各项指标的测定。
2.2.3. 净光合速率测定
净光合速率采用LI-6400便携式光合作用测定系统(美国LI-COR公司生产)测定,在上午9:00~11:00进行,光强控制在1500 µmol∙m−2∙s−1,由仪器配备的红蓝光源(6400-02B LED光源)产生,CO2浓度控制为400 μmol∙mol−1,由小型CO2钢瓶通过仪器配备的CO2注入系统提供,叶面温度分别控制为
30 ℃,
由仪器的加热制冷装置自动精确控制。测定部位为倒二叶,每个材料每株测量1个叶片,共测6个叶片,取平均值作为测定结果。
2.2.4. 表观量子效率与羧化效率测定
表观量子效率(AQY)与羧化效率(CE)也采用LI-6400便携式光合作用测定系统进行测定,于上午9:00~11:00进行,测定部位为倒二叶,每个材料每盆测量1个叶片,共测3个叶片,取平均值作为测定结果。测定时,先通过仪器的Light curve和A-Ci curve自动测量程序获得净光合速率(Pn)对光强(PAR)及同化速率(A)对胞间CO2浓度(Ci)的响应曲线,再根据Pn-PAR及A-Ci响应曲线求出AQY与CE。Light curve自动测量程序的光强控制为2000、1750、1500、750、200、150、100、50、0 µmol∙m−2∙s−1,CO2浓度控制在400 μmol∙mol−1;A-Ci curve自动测量程序的胞外CO2浓度控制为400,300,200,100,50,400,400,600,800,1000,1200,1500,1800 μmol∙mol−1,光强控制在1500 µmol∙m−2∙s−1。两个自动测量程序的叶面温度均控制为
30 ℃
;程序运行前,先让被测叶片在光强为1500 µ mol∙m−2∙s−1下适应15分钟。根据Pn-PAR及A-Ci 曲线数据,以Pn或A为Y值,PAR 或Ci为X值,通过线性回归分析,计算出初始斜率(PAR ≤ 200 µmol∙m−2∙s−1或Ci ≤ 200 μmol∙mol−1),即为AQY或CE。
2.2.5. 光系统II最大光化学量子效率(Fv/Fm)测定
采用LI-6400便携式光合作用测定系统及与其配套的LI-6400-40荧光叶室测定。待测叶片在充分暗适应(30 min)后,通过仪器的叶绿素荧光参数测量程序,测得叶绿素的最小荧光值(Fo)与最大荧光值(Fm),根据Fo与Fm值计算出Fv/Fm,Fv/Fm = (Fm-Fo)/Fm,其中Fv为最大可变荧光。每个材料每株测量1个叶片,共测6个叶片,取平均值作为测定结果。
2.2.6. 叶绿体色素含量的测定
参照王玮 [19] 提取叶绿体色素的方法,取倒二叶中部片段
0.5 克
,用95%的乙醇提取色素,用722型分光光度计测定665 nm、649 nm、470 nm处的吸光度(A),然后根据公式计算出叶绿素(Chl)和类胡萝卜素(Car)的含量,每个材料每株测量1个叶片,共测6个叶片,取平均值作为测定结果。各种色素含量(单位:mg/g)的计算公式为:
公式中V代表提取液总体积(单位:L)。
2.2.7. 数据分析
所得数据采用多重比较法进行差异显著性分析。
3. 结果与分析
3.1. 低温对海南野生稻净光合速率与气孔导度的影响
利用长沙晚秋的自然低温,测定了海南野生稻及供试的2个栽培稻在低温前后的净光合速率与气孔导度。结果表明,在低温胁迫前,海南野生稻的净光合速率及气孔导度与供试的2个栽培稻差异不显著,但低温(6天平均日均温为12.57℃,平均日最低温为9.37℃,平均日最高温为16.60℃)胁迫后,其净光合速率及气孔导度显著高于2个栽培稻对照(图1、图2),且低温胁迫后的值与低温胁迫前的值相比,其净光合速率与气孔导度维持的百分率分别为79.91%与66.53%,也高于2个栽培稻的相应值(分别为69.99%、25.52%与53.16%、21.03%)。说明低温对海南野生稻净光合速率及气孔导度的影响小于供试的栽培稻。
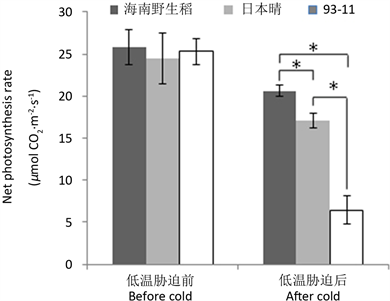 注:“*”表示两者间差异显著(P < 0.05),下同。
注:“*”表示两者间差异显著(P < 0.05),下同。
Figure 1. Effect of low temperature on the net photosynthesis rate in Hainan wild rice
图1. 低温对海南野生稻净光合速率的影响
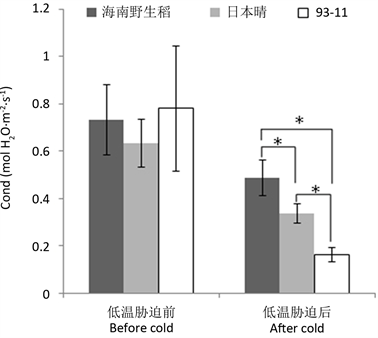
Figure 2. Effect of low temperature on cond in Hainan wild rice
图2. 低温对海南野生稻气孔导度的影响
3.2. 低温对海南野生稻表观量子效率的影响
表观量子效率(AQY)可衡量叶片对照射在其上面的光量子的能量利用效率。对海南野生稻表观量子效率的测定结果表明,在低温前,其AQY与供试的2个栽培稻无显著性差异,但在低温胁迫后,其AQY显著高于2个栽培稻(图3),且低温胁迫后的值与低温胁迫前的值相比,其AQY维持的百分率(64.30%)也高于2个栽培稻对照维持的百分率(34.50%, 18.08%)。说明低温对海南野生稻光能利用效率的影响小于栽培稻。

Figure 3. Effect of low temperature on the AQY in Hainan wild rice
图3. 低温对海南野生稻表观量子效率(AQY)的影响
3.3. 低温对海南野生稻羧化效率的影响
羧化效率(CE)可反映植物叶片活体条件下的羧化酶活性。对海南野生稻羧化效率的测定结果表明,低温胁迫前,海南野生稻的CE与栽培稻差异也不显著,而在低温胁迫后则显著高于2个栽培稻(图4),且低温胁迫后的值与低温胁迫前的值相比,其CE维持的百分率(75.2%)也高于所有栽培稻对照维持的百分率(30.3%, 14.5%)。说明海南野生稻在受低温胁迫后依然保持较高的羧化酶活性。
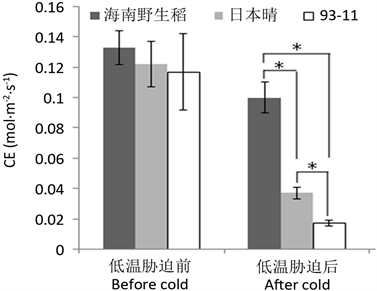
Figure 4. Effect of low temperature on the CE in Hainan wild rice
图4. 低温对海南野生稻羧化效率的影响
3.4. 低温对海南野生稻光系统II最大光化学量子效率的影响
光系统II易受低温、高温等逆境损伤,受损程度可用其最大光化学量子效率(Fv/Fm)来衡量,Fv/Fm反映植物的潜在光合能力。对光系统II的最大光化学量子效率测定结果显示,在低温胁迫前,海南野生稻及2个栽培稻的Fv/Fm值均介于正常范围,且差异不显著,而在低温胁迫后,均低于正常范围,但海南野生稻显著高于所测栽培稻(图5),且低温胁迫后的值与低温胁迫前的值相比,其Fv/Fm值维持的百分率(83.4%)也高于所测栽培稻维持的百分率(67.5%, 38.8%)。说明海南野生稻光系统II对低温的忍耐能力强于2个栽培稻。

Figure 5. Effect of low temperature on the Fv/Fm in Hainan wild rice
图5. 低温对海南野生稻光系统Ⅱ的最大光化学量子效率(Fv/Fm)的影响
3.5. 低温对海南野生稻光合色素含量的影响
对海南野生稻光合色素含量的测定结果表明,在低温胁迫前,其叶绿素a、叶绿素b、总叶绿素及类胡萝卜素含量与2个栽培稻对照差异不显著(表1);低温胁迫后,其叶绿素a、总叶绿素与类胡萝卜素含量均显著高于2个栽培稻对照(表1),且低温胁迫后的值与低温胁迫前的值相比,其叶绿素总量与类胡萝卜素含量维持的百分率也高于所测栽培稻对照维持的百分率(表1)。

Table 1. Effect of low temperature on the contents of photosynthetic pigments in Hainan wild rice (mg∙g−1fw)
表1. 低温对海南野生稻光合色素含量的影响(mg∙g−1fw)
注:Chla、Chlb、ChlT及Car分别代表叶绿素a、叶绿素b、总叶绿素与类胡萝卜素。表中数据为平均值±标准差。同列平均数后带不同小写字母者,差异显著(α < 0.05)。
4. 讨论
4.1. 海南野生稻光合作用对低温的耐性较强,可用于栽培稻的遗传改良
水稻是原产热带或亚热带的一种喜温作物,绝大多数品种的耐冷性不强,尤其是广泛种植的籼稻主栽品种及杂交组合耐冷性不强,限制了其时空分布。耐冷性不强表现在很多方面,其中光合作用对低温的忍耐能力弱是一个重要方面。因此,提高栽培稻光合作用对低温的忍耐能力,是提高其耐冷性的一个重要举措,有助于其充分利用温光资源,拓宽时空分布,从而可大幅提高产量。
野生稻中蕴藏许多优、特异资源,发掘并利用这些资源及其优良性状是对栽培稻进行遗传改良的有效途径。我们对海南野生稻受到低温胁迫后的光合生理特性研究表明,该野生稻在低温胁迫后的净光合速率、叶绿素与类胡萝卜素含量、光系统II最大光化学量子效率(Fv/Fm)、表观量子效率及羧化效率均显著或极显著高于典型栽培稻;而且,与适温下(低温前)测定值相比,上述光合生理指标值维持的百分率也明显高于典型栽培稻。说明海南野生稻光合特性优良。通过育种途径,可望将这些优良光合生理指标性状转入栽培稻,从而提高栽培稻光合作用对低温的忍耐能力,达到拓宽其时空分布、提高产量的目的。
4.2. 该研究加深了对野生稻光合作用的认识
国内外有关野生稻光合作用的研究报道不多,且这些工作主要是研究了适宜环境(顺境)下野生稻的光合特性,而有关非生物逆境对野生稻光合生理特性影响的研究更少。该研究以海南野生稻为材料,探讨了低温对其光合作用的影响,研究结果表明,海南野生稻光合作用对低温的忍耐能力强于典型栽培稻。之所以表现出这一优点,很可能与其低温胁迫后依然能保持较高的叶绿素与类胡萝卜素含量、较高的表观量子效率与表观羧化效率以及光系统II对低温的忍耐能力较强有密切关系,这些具有重要参考价值的数据和结论,无疑加深了我们对野生稻光合作用的认识。
5. 主要结果与结论
经6天平均日均温12.57℃、平均日最低温9.37℃及平均日最高温16.60℃的低温胁迫后,海南野生稻的净光合速率、气孔导度、叶绿体色素含量、光系统II的最大光化学量子效率、表观量子效率及羧化效率均显著高于典型栽培稻;且低温胁迫后的值与低温胁迫前的值相比,其上述光合生理指标值维持的百分率也显著高于栽培稻维持的百分率,表现出其光合作用耐低温的优良特性,可望作为育种资源来提高栽培稻光合作用对低温的忍耐能力。
基金项目
湖南省自然科学基金项目(10JJ3028,2018JJ3343),杂交水稻国家重点实验室(武汉大学)开放基金项目(2016),湖南省重点科技计划项目(2012FJ2013)。