1. 引言
香椿(Toona sinensis Roam)原产于中国,又名香铃子、椿天、香椿芽等,已列入我国珍贵树种名录 [1],成材用时短,树木粗壮高大,材质纹理美观独特,有“中国桃花心木”的佳誉 [2]。香椿适应能力强,分布广泛,较耐湿,喜光照,可在pH 5.5~8.0的土壤环境正常生长 [3]。我国香椿食用历史久远且深受百姓喜欢 [4] [5],有治疗外感风寒、缓解轻度胃痛、润肤明目等功效 [6]。目前,大多数香椿研究集中于集约化栽培 [7] [8] [9]、贮藏保鲜 [10] [11]、化学药理成分等方面 [12] [13],少数机构开展了表型研究 [14] [15],对山东省分布的种质资源表型差异性分析研究较少。本研究以山东主要分布区的香椿为材料,分析种实表型性状间的差异,以对其种质资源收集保护、评价、利用等提供参考。
2. 材料与方法
2.1. 山东省香椿分布情况及资源采集
按照山东省林木种质资源调查结果,香椿在山东各地市均有分布。分组对山东主要分布区11个地市进行采集,共收集资源233份。采集信息见表1。
2.2. 香椿种实表型性状测定
从每份香椿资源中随机选出30粒香椿蒴果,用针叶/种子扫描系统(加拿大WinSEEDLE)对其进行扫描测定。测量蒴果长、种子长(带种翅)、种仁长(去种翅种子),蒴果宽、种子宽、种仁宽。每次测量观察种子的饱满度以及完整度,并对种子长宽进行校准,防止数据出现偏差。种子经双15 (温度15℃,空气湿度15%)干燥至恒重,每份种子随机选出100粒,4次重复,用电子天平称量,精确到0.01,得千粒重。

Table 1. County level planning schedule
表1. 县域等级规划一览表
2.3. 发芽试验
分别选取11地市饱满完整的种子,搓掉种子,浸种完毕后放到光照培养箱(杭州绿博LGZ-380Y)中25℃进行发芽,每单株设置3个重复。每天统一调查发芽势和发芽率,直至发芽结束。
2.4. 数据统计及分析
采用Excel2007进行数据统计整理,采用SPSS18.0进行方差分析、聚类分析、相关性分析和多重比较分析。
3. 结果与分析
3.1. 表型性状差异性分析
对香椿表型性状差异性研究发现,蒴果长度在22.1~29.6 mm之间,排列顺序从短到长依次为QD、JN、DZ、ZB、LY、LC、YT、BZ、HZ、JNI、TA,较长的蒴果分布于HZ、JNI、TA等山东西南部;ZB、LY、LC、YT、BZ、HZ、JNI、TA这8个地市差异不显著,但分别与QD、JN、DZ这3地市差异显著。蒴果宽度在8.9~13.4 cm之间,从短到长依次为QD、BZ、YT、LY、JN、DZ、LC、ZB、HZ、TA、JNI,和蒴果长一样,蒴果较宽的也分布于山东西南部;HZ、TA、JNI均与QD地区蒴果宽差异极显著,与BZ、YT、LY差异显著,与JN、DZ、LC、ZB差异不显著。种子长度在14.0~17.5 mm之间,从短到长依次为JN、DZ、LY、LC、ZB、QD、JNI、HZ、BZ、YT、TA,TA与JN、DZ、LY种子长差异显著,与其他地市的种子长差异不显著。种子宽度在4.5~5.5 mm之间,从短到长依次为LY、JN、QD、DZ、BZ、HZ、YT、LC、JNI、TA、ZB,ZB与LY、JN种子宽差异极显著,与QD、DZ、BZ种子宽差异显著,与HZ、YT、LC、JNI、TA种子宽差异不显著。种仁长度在5.3~6.7 mm之间,从短到长依次为LY、LC、ZB、JN、DZ、BZ、QD、HZ、JNI、TA、YT,YT与LY、LC种仁长差异显著,与其他种仁长差异不显著。种仁宽度在3.6~4.0 mm之间,各地差异不显著。千粒重在8.4~11.9之间,从小到大依次为DZ、BZ、JN、LC、LY、JNI、ZB、YT、QD、TA、HZ,HZ地区的千粒重最大,与DZ、BZ、JN、LC、LY、JNI差异极显著,与ZB地区差异显著,与YT、QD、TA地区差异不显著。比较结果详见表2。
Table 2. Analysis on the differences of phenotypic characters of Toona sinensis
表2. 山东主要分布区香椿种实表型性状差异性分析
注:小写字母代表0.05水平显著;大写字母代表0.01水平显著。
3.2. 表型性状变异分析
山东主要分布区香椿表型性状变异系数见表3。香椿11个群体的各表型性状变异系数范围是7.7%~19.2%,JNI和LY两个群体的变异系数最大,分别为14.9%、14.4%,表明其表型多样性比较丰富;而LC和JN两个群体的变异系数最低,分别为9.4%、9.7%,表明其表型性状变异稳定性较高。香椿11个群体的表型性状变异系数由大至小的排列顺序JNI > LY > TA = ZB > HZ > QD > BZ > DZ = YT > JN > LC,表明山东中南部地区香椿表型多样性程度较高。从香椿6个表型性状的变异系数看,种仁长、蒴果长的平均变异系数分别为13.5%、12.6%,种仁长的平均变异系数明显高于蒴果长的平均变异系数;种仁宽和蒴果宽的平均变异系数分别为11.4%、11.7%,二者相差甚微;表明香椿种仁的稳定性低于其蒴果的稳定性。
Table 3. Variation coefficient of phenotypic traits of Toona sinensis in main distribution areas
表3. 山东主要分布区香椿表型性状变异系数(%)
*表示在0.05水平相关性显著;**表示在0.01水平相关性显著(下同)。
3.3. 表型性状相关性分析
利用SPSS 18.0对香椿6个表型性状进行Pearson相关性分析 [16],结果见表4。如表所示,香椿蒴果长与种子宽呈正相关性,相关系数为0.61;蒴果宽与种子宽、种仁宽呈正相关性,相关系数分别为0.72、0.71;种子长与种仁长呈正相关,相关系数为0.70;种子宽与种仁宽呈极显著正相关,相关系数为0.78;千粒重与种仁长呈正相关,而与种仁宽的相关性不明显,相关系数分别为0.79、0.57。树龄和表型性状间没有直观的相关性。
Table 4. Correlation analysis of phenotypic traits of Toona sinensis in main distribution areas
表4. 山东主要分布区香椿表型性状相关性分析
3.4. 表型性状与生态因子间相关性分析
对山东主要分布区11个香椿群体的表型性状与地理生态因子进行相关性分析,结果如表5所示。香椿蒴果宽与海拔高度呈负相关,相关系数为0.79,表明蒴果宽度随海拔高度增加而变小;种子长与年平均气温呈正相关性,相关系数为0.68,表明种子长度随年平均气温升高而变大。
Table 5. Correlation analysis between phenotypic characters and ecological factors
表5. 香椿表型性状与生态因子间的相关性分析
3.5. 表型性状聚类分析
使用SPSS 18.0统计软件进行聚类分析。聚类方式选择组间平均连接法(Between-Groups Linkage),度量标准采用Euclidean距离,对山东主要分布区11个群体种实表型性状数据进行聚类分析,结果见图1。图中可见11个群体在阈值为10时,可以分成5大类。第一类包括LC、LY、ZB、DZ、JN距离较近,BZ、YT聚为一类,JNI和TA聚为一类,QD、HZ各成一类。JNI、TA、QD等地表型差异显著,而LC与LY,DZ与JN等地表型相似度较高。说明各种源之间的相似程度与地域、气候因子之间呈现不完全相关性。
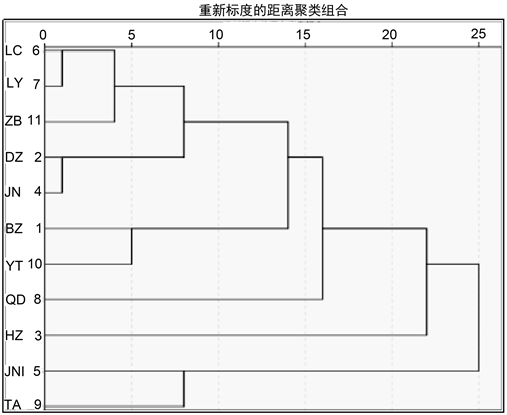
Figure 1. Cluster analysis of Toona sinensis phenotypic traits
图1. 香椿表型性状的聚类分析
3.6. 发芽实验
3.6.1. 发芽势、发芽率和表型性状的相关性
发芽率、发芽势和其表型性状的相关性见表6。结果表明,香椿发芽率与蒴果长呈极显著负相关,发芽势与蒴果长呈显著负相关,说明蒴果越长发芽率和发芽势越低。发芽率、发芽势和其他表型性状未发现直观明显相关性。
Table 6. Correlation between germination potential, germination rate and phenotypic traits
表6. 香椿的发芽势、发芽率和其表型性状的相关性
3.6.2. 发芽势、发芽率和地域因子的相关性
发芽率、发芽势与地域因子的相关性见表7。结果表明,香椿的发芽率与海拔之间表现出一定的相关性,这可能与气候适应有关。发芽率、发芽势与经度、纬度未发现相关性,说明香椿在山东地区属于适生区域,经纬度不是香椿繁育的限制性因子。
Table 7. Correlation between germination potential, germination rate and geographical factors
表7. 香椿的发芽势、发芽率和地域因子的相关性
4. 讨论
植物表型变异是遗传变异的重要表型形式,是基因型和环境因子共同作用的结果,反映了它对于环境因子变化的适应性。本实验对山东主要分布区11个群体进行研究发现,香椿蒴果长宽、种子长宽、种仁长宽和千粒重均存在不同程度差异性。这种差异性应是地理环境因子和自身遗传因子双重作用,并经长时间的自然选择出现的 [17]。从变异分析看,蒴果长、种仁长两性状的变异系数最大、变异程度最高;JNI和LC群体变异系数最大,两个区域表型多样性程度较高,具有较丰富的变异,可对香椿优良品种选育方向提供一定的理论基础。从与地理生态因子的相关性分析看,香椿蒴果长宽、种子宽、种仁长是影响表型性状多样性的重要因素;蒴果宽度、种子宽度皆随海拔高度增加而变小,种子长度随年平均气温升高而变大。种实与经纬度、年降水量之间的相关性暂未发现明显规律,这可能与山东省内各地市间的经纬度差别较小有关,有待进一步研究分析。从聚类分析看,LC与LY、BZ与YT最先聚类,但地域不完全接壤、生态因子也不完全一致,这可能存在地域间的相互引种关系;ZB、DZ、JN地域相连、气候基本一致,可能是形成相似性的原因;QD、HZ、JNI和TA这4个群体各成一类,说明4个群体的香椿资源具一定地域特色,可作为遗传育种资源的重要选择地。从发芽实验看,发芽率、发芽势与蒴果长呈显著负相关,与其他表型性状和地域因子相关性还需进一步探索分析。
基金项目
山东省农业良种工程项目《林木种质资源收集保护与精准鉴定》(2019LZGC018)。
参考文献
NOTES
*通讯作者。