1. 引言
内聚力学说是Dixon和Joly (1894)提出的,尽管从提出到现在一直争议不断,但仍然作为当前解释植物体内水分长距离运输的主流学术观点。其主要原因,就是到目前为止,仍然没有一个可以取代的能够完美解释植物水分传输机制的理论学说。对内聚力学说的质疑,主要是其预测的木质部内的巨大负压和连续水柱至今仍然没有得到直接的实验证据。蒸腾状态下,木质部内会产生大量的空穴已是学术界普遍认可的事实。空穴的发生必然导致水柱断裂,水柱不连续(万贤崇等,2007),就无法靠水的内聚力把水分提升到植物的顶端,因此许多理论和实践方面的问题已无法用内聚力学说做出令人满意的解释。多年以来,人们一直把植物水分运输研究的重点放在断裂的水柱是如何修复上,因为按内聚力学说的观点,水柱不修复水分就不能正常运输。但事实证明,大量的水分运输经常是在水柱不连续的状态下进行的,这说明连续水柱对水分运输并非是必不可少的。由于对根压在蒸腾状态下的作用的低估及对空穴发生机理和消除机制缺乏统一的认识,导致植物水分传输机理方面的研究一直难以取得新的突破。本文通过双渗透泵协同运输模型的建立,提出了连续水柱和不连续水柱的不同移动方式及不连续水柱分段移动的可能性和支持依据。同时对连续水柱是否必须,空穴的发生与消除原理等热点和焦点问题提出了新的见解。
2. 双渗透泵协同运输模型
水分从土壤进入根的中柱是靠渗透作用,叶脉导管中的水分进入叶肉细胞也是靠渗透作用 [1]。通过渗透作用,可以把水势差转变为驱动水分移动的动力。根系的内皮层细胞和叶脉导管至气孔下腔的细胞层都可以看作是一个渗透膜的作用 [2],渗透膜和水势差是发生渗透的基本条件,二者缺一不可。由于通过渗透把水势差变成了水的单向加压或减压作用,所以称之为渗透泵。正是由于根系渗透泵对木质部产生的正压力和叶片渗透泵产生的负压的共同作用,才使水分能够克服重力和摩擦阻力上升到几十米以上的高度。为了解释双渗透泵与木质部压和水分连续状态的关系,用以下模型进行说明。
如图1所示,W1、W2为两个泵(真实模型采用的是流速可调的直流电动隔膜泵),通过管道串联为一个系统。W1的作用是为系统注水加压,W2的作用是为系统吸水减压,两个泵形成的合力使水分沿同一个方向移动。虽然每个泵都能单独工作,但只有两个泵同时工作时,才能使整个系统的传输效率达到最大化。W1的速率为V1,W2的速率为V2,系统压力为P (真实模型用压力真空表观测压力变化)。分别调节W1和W2的流速,当V1大于V2时,系统压力P为正值,驱动力以下面的压力为主,上面的吸力为辅,系统内的水处于充满状态,水柱从上到下是连续的;当V2大于V1时,系统压力P为负值,驱动力以上面的吸力为主,下面的压力为辅,系统内出现水分亏缺,水柱呈断断续续的状态;当V1与V2相等时,系统压力P维持原有的状态不变。
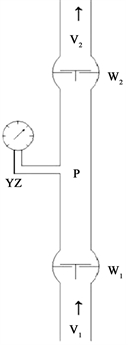 W1加压泵,W2减压泵,V1进水速率,V2出水速率,P系统压力,YZ真空压力表。
W1加压泵,W2减压泵,V1进水速率,V2出水速率,P系统压力,YZ真空压力表。
Figure 1. Sketches of transportation collaboration in double osmotic pump
图1. 双渗透泵协同运输示意图
如果把根系的渗透泵作用比作W1,把叶肉细胞从叶脉导管的渗透吸水看作是W2。V1为根系吸水速率,V2为叶片蒸腾失水速率。木质部作为连接两个泵的管道,那么整个植物的根系吸水和叶片蒸腾失水过程即和以上模型极为相似。当根系吸水速率V1大于叶片蒸腾失水速率V2时,木质部中的压力P为正值,木质部中的水分出现积累,输水通道内的水逐渐饱和,水柱呈连续状态,水分移动的动力以根系渗透泵产生的压力为主。当叶片蒸腾失水速率V2大于根系吸水速率V1时,木质部压力P为负值,输水通道内的水分呈亏缺状态,水柱无法连续,水分移动的动力以叶片蒸腾产生的负压为主。两个渗透泵产生的驱动力是推动水分逆重力方向运行的主要动力,其他方面的影响因素只能起到一定的辅助作用。这一过程中,水势差是唯一的能量来源 [3],渗透膜是能量转换工具,二者缺一不可的,否则,水分的运输就无法持续进行。虽然去掉地上部的根系和去掉根系的枝叶都能独立进行吸水,但要完成大量水分的运输必须靠两个渗透泵形成的合力才能共同完成。如去掉根系的情况下,蒸腾较弱时从切口处直接吸收的水分还能勉强维持体内的水分平衡,但蒸腾强烈的情况下,即使基部切口处供水充足,也仍然不能满足地上部蒸腾的需求,此时如果基部提供一个外加压力的供水装置,萎蔫的叶片马上就会恢复挺立状态。这充分证明,根系的渗透泵作用并不是只有在没有蒸腾或蒸腾微弱的情况下起作用 [4],而是在蒸腾强烈的情况下也仍然发挥着必不可少的重要作用,因为根系渗透吸水产生的正压是木质部正压的唯一来源,无论木质部负压持续多长时间,迟早都是靠根系渗透泵产生的正压来平衡。不论是否有蒸腾发生,只要根系吸水都会对木质部产生正压。即使木质部压力为负值时,也不一定说明根系没有正压产生,只是被蒸腾产生的负压所平衡。所以根压和木质部压应视为两个概念,不能因为木质部压力为负时就认为根系渗透泵作用产生的正压不存在了。干旱季节蒸腾产生的负压经常会持续到夜间(Tyree et al., 1986; Holbrook et al., 2001),所以木质部负压也是根系夜间吸水的主要动力之一。
木质部压力P越高,越有利于水分的向上运输,但是达到一定程度后就会与土壤溶液的水势平衡,使根系内皮层两侧的水势差消失,根系吸水停止。所以无论多高的树木,木质部的正压力一般都不超过0.2 Mpa。木质部压力P越低,与土壤水分之间的水势差就越大,越有利于根系从土壤中吸水,因为正压升高水势,负压降低水势,这同细胞利用膨压吸水的原理是一致的。
蒸腾产生的负压只能传递到根导管末端,而不会直接传递给土壤水分,因为根系的内皮层细胞和皮层细胞对木质部内的流体压力传递起阻挡作用,流体压力的传递只能通过自由间隙和通道而不能通过活细胞传递,所以负压传递到根部后,仍然是通过降低根导管压力势,加大导管与土壤溶液之间的水势差来促进根系的渗透吸水,这也是根系渗透泵作用之所以如此重要的根本原因。
蒸腾不仅降低木质部内的压力势,同时也能使木质部内的渗透势降低,因为叶肉细胞是以渗透方式从叶脉导管内吸水的,蒸腾使大量水分通过叶肉细胞表面散发后,木质部液的水分减少了,但溶质并没有随水分移出,所以蒸腾也会使木质部的渗透势降低 [5]。叶脉导管周围的叶肉细胞虽然没有象根系凯氏带那样的结构,但是叶肉细胞从叶脉导管内吸水必须是以渗透方式进行的。因为只有以渗透方式,才能保证木质部溶液中的各种溶质成分不至于随水分移动到叶肉细胞的表面而丢失。
压力势和渗透势的降低都会为根系吸水提供持续的水势差。因此,双渗透泵协同运输机制的建立,为解决空穴发生及消除、运输动力传输机制等问题奠定了基础。
3. 空穴产生的原因与消除机理
空穴的发生是植物水分运输过程中必然涉及到的一个问题,因为无论是蒸腾状态还是非蒸腾状态,木质部中经常有空穴存在是无可否认的事实。尽管纯水的内聚力理论上可以达到几十Mpa以上,但是木质部液中不仅溶有各种矿物质及有机物,而且还溶有一定的气体,当木质部压力降低到水气分离压时,溶于水中的气体就会以微小气泡的形式逸出,随着压力进一步降低,微小气泡会出现合并、膨大形成空穴 [6],在水分通道内占据一定的空间。把充水正常的一段枝条放入密闭的容器内并对其降压,当压力降低到−0.04 Mpa时就会有气泡逸出,压力越低,逸出的气泡越多。虽然土壤水分在进入根系的过程中经过内皮层细胞的渗透过滤,但是木质部液中仍然溶有一定的气体是无法避免的。当系统中的压力或局部压力由低变高时,有些空穴就会变小或消失 [7],而不需要整个木质部为正压时空穴才会消除(Zimmermann and Brown, 1971; Kramer, 1983)。木质部压力的恢复,主要靠根系渗透泵作用产生的正压来抵消蒸腾产生的负压。当木质部压力由高变低时,空穴由少变多。当压力由低变高时,空穴逐渐减少至全部消失(Sobrado et al., 1992; Edwards et al., 1994)。随着木质部压力的不断变化,空穴的产生和消除、大小和位置都是不断变化的。由此看出,空穴的发生主要是压力降低引起的,当压力恢复到当初的水平时,逸出的气体又会全部溶解到木质部液中 [8]。所以气种是来源木质部液中溶解的气体,然后随着压力的恢复又重新溶解到木质部液中,气体从哪儿来然后又回到哪儿去。这样解释空穴的发生原理和消除机制,既无自相矛盾,又能形成往复循环的机制,与其它学科的理论也没有冲突。如果认为形成空穴的气种是外界气体从纹孔进入导管或管饱,那么气体进入或排出所需要的压力差从哪里来就是个问题(Steu-dle, 2001; Tyree & Zimmermann, 2002)。根据纹孔的结构,正常情况下水分子穿透纹孔膜是比较容易的,但气体分子穿过纹孔膜是相当困难的,从这一点来看,外界气体进入导管或管饱的可能性不大,反而为解释空穴的消除机制设下了一个难题。
空穴的发生和存在,还可以看作是木质部水分亏缺的一种状态,因为在蒸腾强烈的夏季,木质部中经常有50%的水被空穴代替(Kramer, 1983)。当木质部为负压时,导管和管饱的径向收缩是非常有限度的,水在常温下又是不可膨胀的,在水分不能充满所有管腔的空间时,就必然有空隙存在,这些空隙就是由逸出的气体在负压作用下膨胀形成的空穴所占据。如果木质部水分始终是饱和的,那么也就不会有空穴存在的可能。所以这也是根系吸水不能满足蒸腾失水的必然结果,只有当根系吸水速率大于蒸腾速率时,才能将亏缺的水分补足,空穴也即随之消失。
栓塞从概念上与空穴应有所区分,栓塞应该定义为外界空气进入导管或管饱形成的。空气通过伤口在负压作用下吸入导管或管饱形成的栓塞,能起到隔离和保护伤口的作用,但外界空气一旦进入到正常的输水通道内,则会严重阻碍水分的运输,而且靠植物自身很难再排出到体外或在体内溶解消除,这种情况只有在管壁失水严重纹孔结构发生变化时才会有可能出现,对植物会造成不可复性的损害。空穴发生后并不妨碍水分的输送(Wagner et al., 2000),但栓塞则会严重阻碍水分的正常通行。所以只有把空穴和栓塞区别对待,才能更有利于对空穴和栓塞的正确解释。
4. 木质部水分的分段移动及支撑依据
蒸腾状态下木质部内水柱不连续可以说是一种常态。事实证明,木质部在水柱不连续的状态下水分运输并没有停止而是仍然在继续(Zimmermann et al., 2004)。那么在水柱不连续的情况下是如何进行运输的就成为需要研究解决的一个关键问题。许多研究已经证明,叶片蒸腾产生的吸水动力传递到根部需要相当长的时间 [9],从这一点即可以肯定,蒸腾产生的吸水动力并不是靠内聚力传递的,无论水柱是直线连续还是曲线连续,从传递速度和时间上看,靠内聚力传递蒸腾拉力都是不可能的。假如水的内聚力达到30 Mpa,那么水柱的抗张强度可达到铜线的三分之一,这种强度近似固体的水柱传递拉力,无论树干多高,从叶片到根部不会超过几秒钟的时间,根本就不需要几十分钟甚至几个小时。所以,用连续水柱来解释蒸腾拉力的传递是难以成立的。
木质部的水分运输管道可以看作是一束上下相通、纵横相连(通过纹孔横向连通)的毛细管,如果毛细管中从上到下全部充满水分(即水柱呈连续状态),所有管腔内只有液相而没有气相存在,也就不会有毛管力产生,系统内由重力所产生的压力与普通管道内是相同的。当空穴产生后,连续水柱被分成若干段,每一段的上部都有一个气液交界的凹面,每一个凹面产生的毛管力支撑一段水柱,从根部导管到叶脉导管之间,由若干个凹面就可以把每段水柱固定在管腔内,靠毛管力与重力平衡,在没有压力梯度作用情况下,每一段水柱既不向上移动,也不向下移动,就像一级一级的阀门将每一段水柱托住而不会因重力作用而下移,所以无论多高的树木,蒸腾状态下在树干基部均测不到由重力产生的0.01 Mpa/m,的压力,整个木质部内的任何部位也都不需要0.01 Mpa/m,的压力来支撑,这也是水分能够实现分段移动的一个重要的前提条件。当有压力梯度存在时,每一段水柱,都可以在压力梯度的驱动下单独移动,而无需上下一起连动。
根据双渗透泵协同运输原理,植物体内的水分只有在根系吸水大于蒸腾的情况下才会以连续水柱的方式运行,也就是说木质部为正压水柱才能保持连续状态,而连续水柱的驱动力是以根系产生的向上的压力为主而不是叶片蒸腾产生的拉力为主,所以水柱几乎不需要承受张力,也就不存在靠内聚力传递蒸腾拉力的问题。蒸腾初期消耗的水分主要是上部的枝干内贮存的水分 [10],包括导管和管饱及环管薄壁细胞内的水分,所以并不是一有蒸腾就马上就传递到根系吸水。蒸腾产生的负压首先发生在叶脉导管和小枝内,然后随着水流依次向上移动,负压向下传递,一直到根的中柱导管。根的渗透泵作用产生的正压力,会持续推动水分向顶部的低压区移动。根据实验证明,负压的传递速度与水分的移动速度是基本一致的,一般在0.45~50 m/h。所以这是一个压力平衡过程,水分的移动是一节一节地在压力梯度作用下蠕动式运行的,而不是靠连续水柱把整个木质部的水一起拖拽上去。这种分级移动的方式只需要较小的压力梯度就能驱动被分段的水柱。当空穴存在时,如果空穴下面的压力高于空穴的压力时,水分即向上移动,空穴就会变小或消除。下部的压力降低后也会出现新的空穴。相邻的导管之间,水分也会在压力梯度作用下通过纹孔进入到有空穴的部位。水分也可绕过空穴通过纹孔进入到相邻的导管内向上运行。在同一部位空穴的发生与消除会交替进行,原有的消除了,可能还会有新的再产生,所以空穴的产生与消除是随着压力的变化而不断变化的,水分的上升路径也是曲曲折折向上运行的。
由以上分析可以看出,实现分段移动必须具备以下条件:一是水柱的任意一处随时都可以发生断裂,压力恢复后又可以迅速恢复;二是每段水柱都必须能够单独移动,而不是整条水柱从上到下一起移动;三是每段水柱都必须有一种力与重力平衡,使整个木质部内的水分不需要0.01 Mpa/m的压力梯度来支撑,就如同靠多级阀门控制住水分不向下移动的作用是相似的。
5. 与其它理论学说、假说的关系
内聚力学说肯定了太阳辐射对植物蒸腾的作用,这一点与渗透泵原理是一致的,因为是太阳辐射的能量使叶肉细胞的水分不断地蒸发到大气中,使叶肉细胞与叶脉导管之间维持一个持续的水势差,通过渗透,把水势差转化成水分移动的驱动力,而不需要消耗植物自身的代谢能量。但是内聚力学说预测的巨大负压和连续水柱很难得到证实,特别对水柱不连续状态下的运输机理更是无法做出合理的解释。通过连体和离体两种方式测定,蒸腾状态下,木质部内的负压最大也只能测到−0.08 Mpa,这与物理学原理也是一致的,因为地球大气中,负压的最大值理论上不会超过−0.1 Mpa [11],Zimmermann等人用木质部压力探针测得数据也是在0~0.1 Mpa范围内,因此木质部内只存在有限的负压与相关理论观点是一致的(Smith, 1994; Canny, 1995; Zimmer-mann et al., 2004)。如果水柱不连续并且每段水柱靠毛管力与自身的重力平衡,那么也就不存在内聚力大于抗张力的说法。水门(Watergates)假说实际上已经发现了水分阶梯式分段移动的现象,应该说这是一个非常重要的发现,已经触及到解决问题的边缘了,但是分段移动的前提是,导管和管饱中必须要有能够开启/关闭的阀门控制着水分一级一级向上提升,然而,至今始终没有人能够从解刨学上证实导管或管饱中这种“阀门”的存在,虽然发现了这种现象,但未找到其真正的机制,所以该假说也因缺乏有力的证据支撑而没有得到广泛的认可。补偿压学说(Canny)的观点是强调环管薄壁细胞和射线细胞的组织压施加给输导组织中的封闭水柱,使输导组织内的压力梯度得到补偿 [12],但是这种薄壁细胞产生的压力是暂时的,难以作为水柱稳定上升的持续动力。当导管和管饱内的水势低于周围活细胞水势时,活细胞内的水分就会以渗透方式进入到导管内,在短时间内对补充导管水分不足和消除空穴可能会有一定的作用。当导管内水分相对充足时,水势高于活细胞的水势,导管内的水又可以补充到细胞内。所以活细胞的作用主要是起贮存和调节作用 [13]。在没有叶片蒸腾的情况下,只要活细胞(包括芽、叶、果实的活细胞)有足够低的水势,导管中的水分通过渗透作用进入活细胞,对导管水分的上升也会产生一定的拉动作用。所以,活细胞与输水通道之间发生的水分交换也是以渗透方式进行的,符合渗透泵原理,与双渗透泵协同运输机制和分段移动均不矛盾。
从以上研究分析可以看出,空穴的实质是水分亏缺造成的,是随压力降低而产生,随压力升高而消除,空穴的存在会增加水分运输的阻力,但不会造成水分运输停止。连续水柱和不连续水柱是水分运行的两种方式,不是水分能否正常运输的决定因素。水分的分段阶梯式移动,既不需要木质部内有巨大的负压,也不依赖连续水柱,而是利用较小的压力梯度把水柱一段一段逐级提上去。因此,下一步水分生理研究的重点和方向,应放在不连续水柱的运输机制及支撑条件上,而无需再把空穴的消除机制和连续水柱的恢复机理作为植物水分传输机制的焦点和热点问题。