1. 引言
报春苣苔(Primulina tabacum)属于苦苣苔科(Gesneriaceae)报春苣苔属(Primulina)植物 [1] [2]。是国家I级重点保护野生植物 [3]。报春苣苔一般生长在石灰岩山地林下的石上、洞穴入口甚至洞中或河流两岸石壁,为多年草本;其叶为基生叶;伞状聚伞花序;花序有花3~9朵,具对生苞片;高脚碟状花冠 [2]。报春苣苔属植物仅分布在广东北部乳源、连县、阳山以及江西婺源、湖南宁远、广西贺州一带 [4] - [10]。
报春苣苔主要生长在海拔约300 m的石灰岩山洞口附近,在其附近的植物群落主要是一些耐阴湿及喜钙的植物,并有苔藓植物 [11] 伴生。报春苣苔由于生长环境的影响,光源以及大气环境成为了限制其生长范围的重要因素 [12]。近年,人类的活动对报春苣苔生存的环境产生了不利的影响,其种群数量和分布范围也因此面临骤减的危险。对濒危植物报春苣苔的研究有不少,研究的内容涉及生态生物学特征 [12],光合作用特性 [13],花粉形态 [14] 和叶片结构 [15] 等方面,这些研究都表明:现今作为保护植物的报春苣苔越来越受到学者们的重视。目前,关于苦苣苔科一些属种的胚胎学研究仅见少数的报道 [16] [17]。本研究旨在对报春苣苔的雌配子体发育进行解剖学研究,一方面为报春苣苔的保护措施的制定提供有参考性的资料;另一方面也为该属的生殖生物学、发育生物学方面的研究积累资料。
2. 材料与方法
2.1. 材料
研究的不同发育阶段的报春苣苔花,采于广东省连州市星子镇上柏场。用F.A.A固定液(70%酒精90份,冰醋酸5份,甲醛5份)固定,70%酒精低温保存,备用。
2.2. 方法
取固定后的报春苣苔花,用爱氏苏木精整体染色,常规石蜡切片法切片,切片厚度为8 µm,对子房用1%碱性品红酒精液复染,后用中性树胶封固;用Olympus显微镜观察并摄影。
3. 观察结果
3.1. 大孢子发育
在显微镜下观察胚珠纵切面发现:报春苣苔的子房为二心皮一心室,侧膜胎座,心室中含有众多胚珠(图1)。胚珠呈倒生型,薄珠心,珠心外为单层珠被(图2)。孢原细胞起源于珠心表皮下方的细胞,并在以后直接发育成大孢子母细胞。大孢子母细胞的形态结构与珠心的其它细胞明显不一样,体积较大、细胞核大且原生质体丰富(图3)。大孢子母细胞通过减数分裂形成呈直线排列的两个细胞,随后两个细胞再次进行分裂,形成一个直排的四分体(图4)。接着,靠近珠孔端的三个大孢子逐渐出现退化现象,后完全消失;靠近合点端的大孢子继续发育,其体积逐渐增大,进一步发育为功能大孢子(图5);与此同时,珠心和珠柄一侧的细胞分裂速度加快,使珠柄的发育方向发生改变,致使胚珠倒向生长,形成倒生胚珠。珠心两边表皮下的细胞经过分裂,一直向着珠孔端的方向生长,逐渐包住珠心,随后发育成为具有两到三层细胞的单层珠被。因此,其胚珠类型为倒生、薄珠心。
经观察,报春苣苔的大孢子发育阶段在减数分裂等过程存在一些败育现象,如:减数分裂不规则,形成非直线排列的几个大孢子(图6);功能大孢子解体(图7)等。
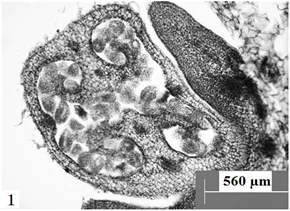
Figure 1. Transverse section of ovary, showing parieta placenta. Scale bars = 560 µm
图1. 子房横切,示侧膜胎座。比例尺 = 560 µm

Figure 2. Ovules longitudinal section, the arrow shows tenuinucellate ovule. Scale bars = 80 µm
图2. 胚珠纵切,箭头所示薄珠心。比例尺 = 80 µm
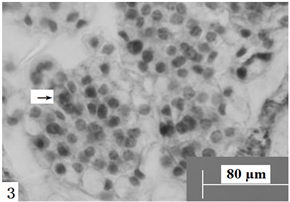
Figure 3. Ovules longitudinal section, the arrow shows megaspore mother cells. Scale bars = 80 µm
图3. 胚珠纵切,箭头所示大孢子母细胞。比例尺 = 80 µm
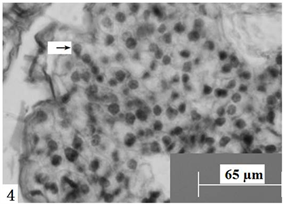
Figure 4. Ovules longitudinal section, the arrow shows four megaspores arranged in a straight line. Scale bars = 65 µm
图4. 胚珠纵切,箭头所示直线排列的四个大孢子。比例尺 = 65 µm

Figure 5. Ovules longitudinal section, the arrow shows functional megaspore. Scale bars = 80 µm
图5. 胚珠纵切,箭头所示功能大孢子。比例尺 = 80 µm

Figure 6. Ovules longitudinal section, the arrow shows megaspores arranged in a “T” shape. Scale bars = 560 µm
图6. 胚珠纵切,箭头所示呈“T”字形排列的大孢子。比例尺 = 560 µm

Figure 7. Ovules longitudinal section, the arrow shows disintegrate megaspore. Scale bars = 90 µm
图7. 胚珠纵切,箭头所示解体功能大孢子。比例尺 = 90 µm
3.2. 雌配子体发育
在显微镜下观察胚珠纵切面发现:报春苣苔胚珠的近合点端的功能大孢子的发育,其体积慢慢增大,细胞质液泡化慢慢加强,胚囊体积逐步增大,功能大孢子发育成单核胚囊(图8),单核胚囊中的细胞核位于胚囊中央。单核胚囊通过一次有丝分裂形成两个细胞核,两个细胞核处于胚囊中央,随后移动到胚囊两侧,即一个位于胚囊的珠孔端,另一个位于胚囊的合点端。同时,胚囊的液泡逐渐增大,在胚囊中央形成一大液泡,此时两个细胞核分别位于胚囊的两端,形成二核胚囊(图9)。二核胚囊继续发育,体积逐渐增大,两个细胞核再分别进行一次有丝分裂,两个细胞核的分裂基本是同步进行的。此时,珠孔端和合点端各有两个细胞核,形成四核胚囊(图10)。位于胚囊两端的四个细胞核再分别进行一次有丝分裂产生八核胚囊(图11),胚囊继续沿着纵轴方向逐渐延长。随后,近珠孔端的一个细胞核向中央移动,近合点端的一个细胞核也向中央移动,后停留在胚囊的中央位置,构成两个并列的细胞核。八核胚囊阶段形成后经过短暂的时间,便进行细胞分化。近珠孔端的三个细胞核,分别形成细胞壁,其中两个细胞核分化成助细胞,另外一个细胞核分化成卵细胞,刚分化的卵细胞的细胞核较大且原生质体丰富。在以后发育中,卵细胞的细胞质中开始出现液泡,液泡化逐渐加强,细胞极性逐渐明显,并细胞形状发生改变,最终呈梨形,其细胞核远离珠孔端。两个助细胞分布于卵细胞的两侧,形成卵器(图12)。近合点端的三个细胞核之间逐渐形成细胞壁,进而分化成三个反足细胞,呈“品”字排列(图13)。反足细胞存在的时间比较短暂,一般在受精前开始退化。位于胚囊中央的两个细胞核结合形成中央细胞,为两核的初生极核(图14)。随着胚囊细胞化的完成,雌配子体形成,进入成熟胚囊阶段。成熟胚囊由单孢子发育形成,具八核七细胞结构;因此,成熟胚囊的类型为蓼型胚囊。

Figure 8. Ovules longitudinal section, the arrow shows mononuclear embryo sac. Scale bars = 200 µm
图8. 胚珠纵切,箭头所示单核胚囊。比例尺 = 200 µm

Figure 9. Ovules longitudinal section, the arrow shows two-nuclear embryo sac. Scale bars = 400 µm
图9. 胚珠纵切,箭头所示二核胚囊。比例尺 = 400 µm
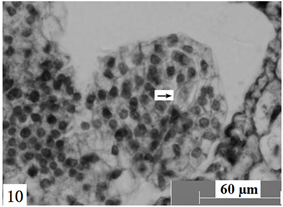
Figure 10. Ovules longitudinal section, the arrow shows four-nuclear embryo sac. Scale bars = 60 µm
图10. 胚珠纵切,箭头所示四核胚囊。比例尺 = 60 µm

Figure 11. Ovules longitudinal section, the arrow shows eight-nuclear embryo sac. Scale bars = 460 µm
图11. 胚珠纵切,箭头所示八核胚囊。比例尺 = 460 µm

Figure 12. Ovules longitudinal section, the arrow shows egg apparatus. Scale bars = 350 µm
图12. 胚珠纵切,箭头所示卵器。比例尺 = 350 µm
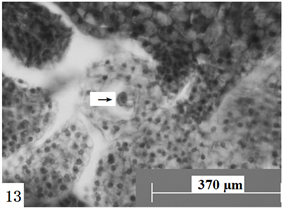
Figure 13. Ovules longitudinal section, the arrow shows antipodal cells. Scale bars = 370 µm
图13. 胚珠纵切,箭头所示反足细胞。比例尺 = 370 µm

Figure 14. Ovules longitudinal section, the arrow shows central cell. Scale bars = 240 µm
图14. 胚珠纵切,箭头所示中央细胞。比例尺 = 240 µm
经观察,在报春苣苔雌配子体发育过程并非所有胚珠都能发育形成成熟胚囊,不少胚珠在不同发育阶段,出现败育的现象,如:单核胚囊解体(图15),形成空泡化的细胞核;二核胚囊形成后,细胞核形成颗粒状结构(图16);四核胚囊形成后,会出现有多种败育现象:四个细胞核解体(图17),或四个细胞核没有分布在胚囊两端,而聚合在一端或中央并形成多核合胞体(图18~20)等。

Figure 15. Ovules longitudinal section, the arrow shows vacuolated mononuclear embryo sac. Scale bars = 60 µm
图15. 胚珠纵切,箭头所示空泡化的单核胚囊。比例尺 = 60 µm

Figure 16. Ovules longitudinal section, the arrow shows disintegrated two-nuclear embryo sac. Scale bars = 90 µm
图16. 胚珠纵切,箭头所示解体的二核胚囊。比例尺 = 90 µm
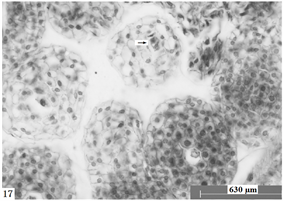
Figure 17. Ovules longitudinal section, the arrow shows disintegrated four-nuclear embryo sac. Scale bars = 630 µm
图17. 胚珠纵切,箭头所示解体的四核胚囊。比例尺 = 630 µm

Figure 18. Ovules longitudinal section, the arrow shows polypolymeric cells clustered at the end of the pore. Scale bars = 410 µm
图18. 胚珠纵切,箭头所示聚在珠孔端的多聚合胞体。比例尺 = 410 µm

Figure 19. Ovules longitudinal section, the arrow shows a polymorphic body clustered in the center of the embryo sac. Scale bars = 400 µm
图19. 胚珠纵切,箭头所示聚在胚囊中央的多聚合胞体。比例尺 = 400 µm
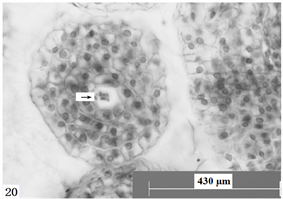
Figure 20. Ovules longitudinal section, the arrow shows a polymorphic body clustered in the center of the embryo sac. Scale bars = 430 µm
图20. 胚珠纵切,箭头所示聚在胚囊中央的多聚合胞体。比例尺 = 430 µm
3.3. 受精及胚的发育
在显微镜下观察胚珠切面发现:报春苣苔的精子细胞分别进入卵细胞与中央细胞(图21、图22),与它们完成受精;合子形成以后经过分裂形成两个细胞,分别是近合点端的顶细胞和近珠孔端的基细胞。基细胞只进行横分裂,经过多次的横分裂后形成极短、细胞少的胚柄,胚柄随后将胚体推到胚囊腔中。顶细胞通过多次分裂形成多细胞原胚,原胚细胞再进行多向分裂形成球形胚(图23),同时胚囊体积逐步增大。随着胚体细胞的继续分裂,球形胚的体积逐渐增大;此时,细胞的分裂方向、频率出现变化,发育成心形胚(图24)、鱼雷胚、子叶胚(图25)。胚囊处于心形胚阶段时,胚柄仍然存在(图26),到子叶胚阶段时,胚柄基本上整体消失。胚乳核分裂较合子分裂要早(图27)。
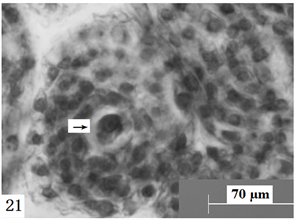
Figure 21. Ovules longitudinal section, the arrow shows sperm and egg apparatus cells. Scale bars = 70 µm
图21. 胚珠纵切,箭头所示精子与卵器细胞。比例尺 = 70 µm
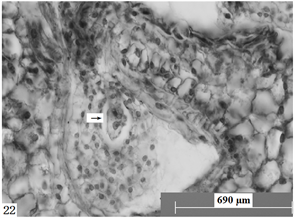
Figure 22. Ovules longitudinal section, the arrow shows sperm, egg cell, central cell. Scale bars = 690 µm
图22. 胚珠纵切,箭头所示精子与卵细胞、中央细胞。比例尺 = 690 µm

Figure 23. Ovules longitudinal section, the arrow shows globular embryo. Scale bars = 250 µm
图23. 胚珠纵切,箭头所示球形胚。比例尺 = 250 µm
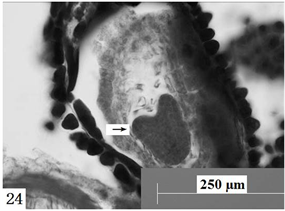
Figure 24. Ovules longitudinal section, the arrow shows heart shaped embryo. Scale bars = 250 µm
图24. 胚珠纵切,箭头所示心形胚。比例尺 = 250 µm
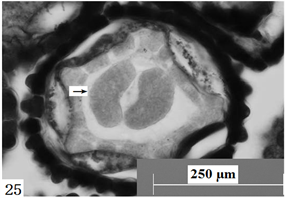
Figure 25. Ovules longitudinal section, the arrow shows cotyledon embryo. Scale bars = 250 µm
图25. 胚珠纵切,箭头所示子叶胚。比例尺 = 250 µm
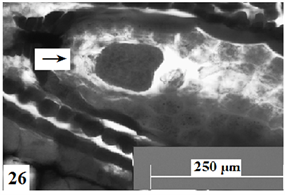
Figure 26. Ovules longitudinal section, the arrow shows a heart-shaped embryo with a suspensor. Scale bars = 250 µm
图26. 胚珠纵切,箭头所示具胚柄的心形胚。比例尺 = 250 µm

Figure 27. Ovules longitudinal section, the arrow shows endosperm nuclei and zygotes dividing. Scale bars = 900 µm
图27. 胚珠横切,箭头所示正在分裂的胚乳细胞核及合子。比例尺 = 900 µm
在观察中发现,在胚囊的发育阶段出现有双胚的现象(图28)。
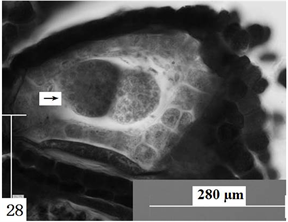
Figure 28. Ovules longitudinal section, the arrow shows embryo sac with double embryo. Scale bars = 280 µm
图28. 胚珠纵切,箭头所示具双胚的胚囊。比例尺 = 280 µm
4. 讨论
从观察结果可以看出:报春苣苔在大孢子发育以及雌配子体形成的各个阶段,都出现不同方式的败育,导致大多数胚珠都不能发育形成成熟胚囊。因此,报春苣苔虽有数量较多的胚珠,但只有少数胚珠能够发育形成成熟胚囊,这就大大减少了该种群的繁殖能力,影响其有性生殖的有效性,因而导致其后代数量难以增加 [18]。由于雌、雄配子体(另文表述)在发育各个阶段都有败育现象发生,因此,该物种自身繁衍能力不强,是其成为保护植物的原因之一。
大孢子以及雌配子体发育异常现象在一些濒危植物的胚胎学研究也有提到,例如:西畴含笑(Michelia coriacea) [19]、珙桐(Davidia involucrate) [20] 均有观察到大孢子以及雌配子体发育异常现象。植物有性生殖的每一个环节都是非常关键的,但在任何一个环节中出现了问题,都有可能致使其生殖出现障碍,进而导致该物种的濒危 [21] [22]。有很多因素可以引起植物出现濒危情况,譬如:存活能力、生殖能力、对环境的适应能力等内在因素。而种群生活史中多个环节都出现问题,便大大加剧了导致个体出现生长发育不良和早期衰退的问题的概率 [23]。导致报春苣苔濒危的因素有很多种,但其生殖过程中出现了因发育异常原因导致的生殖障碍,应该是导致其濒危的重要原因之一。
基金项目
2019年广东省本科高校教学质量与教学改革工程项目:生物科学(特色专业立项项目)。
NOTES
*通讯作者。