摘要:
以峨眉含笑(Michelia wilsonii Finet et Gagnep.)种子萌发后的根尖为实验材料,用DAPI染色法制作染色体标本分析其染色体核型。结果表明,峨眉含笑为二倍体,染色体数目38条,核型公式为2n = 38 = 32m + 4sm + 2st,核型不对称系数为56.3%,核型属Stebbins的2A型,在进化上处于原始的地位。本研究确定了峨眉含笑的核型及进化地位,为其开发利用和保护提供了细胞学资料。
Abstract:
The root tips of Michelia wilsonii Finet et Gagnep. was used for experiment to analyze chromo-some karyotypes by DAPI staining method. The results show that Michelia wilsonii is diploid, there are 38 chromosomes, its karyotype formula is 2n = 38 = 32m + 4sm + 2st. The karyotype asymmetry coefficient is 56.3%, and the karyotype is Stebbins’s 2A, which is in a primordial po-sition in evolution. The study determined the karyotype and evolutionary status of Michelia wilsonii, and provided cytological data for its development, utilization and protection of the en-dangered species.
1. 引言
植物染色体核型分析是将细胞染色体的数目、形态、长度、着丝点位置等内容进行分析研究,是分类和遗传研究的重要手段 [1] [2]。含笑属(Michelia Linn.)全世界约60种,我国约有35种 [3]。含笑属的核型分析,李秀兰、陈瑞阳、孟爱平 [4] [5] [6] [7] 均有报道。峨眉含笑(Michelia wilsonii Finet et Gagnep.)为木兰科含笑属常绿乔木,中国特有,自然种群极小,被列为国家二级保护植物 [8]。自然种群分布范围狭窄,峨眉山仅分布在海拔1000~1300 m的常绿阔叶林中 [9]。该树种树形优美,树干笔直,花重瓣黄色,有芳香,是园林绿化的重要观赏树种和珍贵用材树种 [10]。本研究利用峨眉含笑的种子初生根尖为材料,进行染色体核型研究,旨在分析峨眉含笑的染色体数目和核型类型,为峨眉含笑进化位置的探讨和进一步开发利用和保护等工作的开展提供细胞学资料。
2. 材料与方法
2.1. 材料
本研究供试材料来源于中国西南的峨眉山,采集地海拔1050 m,东经103˚22',北纬29˚35'。9月底待峨眉含笑果实中鲜红色的种粒开始暴露时采收,去掉蓇葖,清洗掉红色种皮,水沉法选取籽粒饱满的种子,阴干备用。
2.2. 方法
1) 种子萌发
用1000 mg/L的赤霉素浸泡峨眉含笑种子24 h后放入人工气候箱,在恒温25℃、相对湿度75%、光照强度6000~8000 lux、光照时间12 h/d的条件下进行萌发。
2) 取根与固定
根长至0.5~2 cm时取样,放入0.5 ml EP管中置于冰上,在其盖上戳一个直径约为3 mm左右的小洞,将EP管放入0.9 Mpa笑气罐中处理2 h,取出EP管置于冰上加入90%乙酸,固定根尖8~10 min [11]。
3) 制片与染色
切下长度约0.5 cm的根尖部分置于果胶纤维素酶混合酶液中,37℃酶解1 h。用70%乙醇洗尽酶液,捣碎根尖,4000 rmp离心2 min,加入15~50 μl乙酸,获得根尖细胞悬浮液,滴片。自然风干后DAPI染色盖上盖片 [12],在Leica荧光显微镜DM2500下镜检。
4) 染色体检测与核型分析
选择分散良好、着丝点清晰的染色体分裂相,对其进行形态观察和数目统计,用GIMP软件分别测量染色体的总长、长臂、短臂。体细胞分裂中期核型分析参照李懋学和陈瑞阳 [13] 的标准,用二点四区系统标准确定着丝粒位置 [14],核型分类按Stebbins [15] 的标准,核型不对称系数参照Arano [16] 的方法。
3. 结果
3.1. 染色体形态及数目
观察了处于有丝分裂中期、中后期、后期细胞分裂相图片,均未观察到随体和端部着丝粒染色体,染色体形态和结构均比较稳定(图1)。选取分散良好、形态清晰的有丝分裂中后期图,从着丝粒个数可以确定,峨眉含笑染色体数目为38条,且为二倍体,故2n = 38 (如图2)。
 (a) 有丝分裂中期
(a) 有丝分裂中期 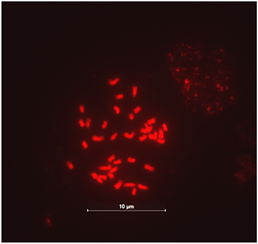 (b) 有丝分裂后期
(b) 有丝分裂后期
Figure 1. Chromosomes morphology of Michelia wilsonii
图1. 峨眉含笑染色体形态

Figure 2. Karyogram of Michelia wilsonii
图2. 峨眉含笑染色体核型
3.2. 核型参数
结果表明(见表1、图2、图3),19对峨眉含笑染色体中,中部着丝粒染色体(m)有16对(第1、3~6、9~19号),近中部着丝粒染色体(sm)有2对(第7~8号),近端部着丝粒染色体(st) 1对(第2号)。核型公式为2n = 2x = 38 = 32m + 4sm + 2st。
基于植物核型分析标准,计算结果(表1)显示,峨眉含笑染色体绝对变异幅度为0.94~1.7 μm,相对长度为4~7.07%,臂比值在1.03~3.62之间,臂比值大于2:1的占10.5%,核型不对称系数为56.3%。最长与最短染色体的比值为1.768。核型不对称性为2A型。

Table 1. The chromosome parameters of Michelia wilsonii
表1. 峨眉含笑的染色体参数
m: metacentric的缩写,表示具中部着丝点染色体;sm: submetacentric的缩写,表示具亚中部着丝点染色体;st: subterminal的缩写,表示近端部着丝点染色体。
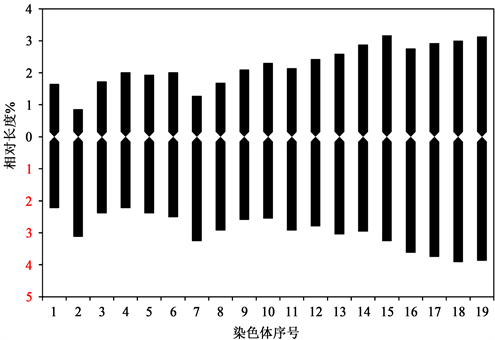
Figure 3. Karyotype pattern of Michelia wilsonii
图3. 峨眉含笑染色体核型模式图
4. 讨论
本研究中峨眉含笑染色体数目2n = 38,为二倍体,与前人报道的木兰科大多数属的染色体数目和倍性一致 [5] [6] [16] - [21]。核型分类为2A型,属于较整齐的对称核型,表明峨眉含笑遗传性状稳定、在进化上处于比较原始的地位,支持了“有花植物中木兰科是最原始的科之一” [7] 这一观点。
李秀兰报道的峨眉含笑的核型公式为2n = 30m + 8sm (2SAT) [4],本报道中峨眉含笑未发现随体,核型公式为2n = 2x = 38 = 32m + 4sm + 2st。造成核型不同的原因,可能是人为的不同实验操作 [22] 影响了染色体的观察,即使是同一个物种,不同的人也有不同的研究结果,李秀兰 [4] 发现深山含笑核型公式为2n = 38 = 32m + 6sm,核型为2A型,而孟爱平 [7] 发现深山含笑核型公式2n = 38 = 28m + 10sm,核型分类为2B型,核型公式和核型分类均不同;同一物种核型不同也可能是物种不同来源地导致了这种差异,有研究证实不同产地的物种,其核型对称性、核型公式、随体有无,存在细微差异 [23],本研究结果佐证了这一观点。可能是物种在进化的进程中,为了适应环境而使染色体变异,最终表现为物种内部染色体核型多态性 [24]。
基金项目
四川省科技计划重点研发项目“峨眉3种濒危植物野生种群自然更新能力评估与人工促进措施”(2019YFS0459)、四川省植物资源共享平台建设项目(2019JDPT0028)和峨眉山植物园科普教育基地能力提升创新与示范(2018KZ0006)。
NOTES
*通讯作者。