1. 引言
肥胖是由于脂肪的过度累积而造成的一种病理状况 [1] 。根据世界卫生组织(WHO) 2014年的调查报告结果显示,自1980年以来,肥胖人群的数量已经接近过去的两倍 [2] 。肥胖是一种慢性、复发性的疾病,其特征在于人体脂质代谢紊乱和能量失衡,严重危害人类健康,并与糖尿病、动脉粥样硬化等代谢性疾病密切相关 [3] 。研究表明,肥胖机体存在慢性低水平的炎症状态,而Toll样受体(Toll-like receptors, TLRs)在这种低度炎症状态的形成中起到重要作用,并且影响机体的物质代谢。尽管肥胖机体的炎症反应没有典型的红肿热痛表现,但其具有经典炎症反应相似的炎性分子和相关信号传导通路参与,而这种持续存在的炎症状态会导致代谢紊乱和功能障碍 [4] 。在肥胖患者中,TLRs被内源性配体激活,从而参与机体的炎症反应的产生 [5] 。在高脂饮食条件下,脂肪酸和脂多糖(Lipopolysaccharide, LPS)可以促进巨噬细胞、脂肪细胞和骨骼肌细胞表达TLR2/4及其下游信号通路,从而引发炎症反应。TLR6在与肥胖相关的慢性炎症状态和病态肥胖患者的肝脏疾病中也发挥着重要作用,在非酒精脂肪肝的患者中,TLR6在外周血单核细胞和肝脏中的表达明显增高 [6] 。然而,TLR6水平的变化与肥胖、机体低度炎症水平的关系如何还不清楚,是否是因高脂饮食引起TLR6的变化导致机体低度炎症水平继而引起肥胖的发生也不明了。因此,TLR6所参与的信号通路,与肥胖患者的炎症反应的机制需要进一步的研究。已知TLR6与TLR2形成异质二聚体,并参与炎症信号的传递,其激活依赖于MyD88蛋白(myeloid differentiation factor 88),从而引发一系列炎性因子的表达 [7] 。MyD88是TLR信号转导途径中的主要适配蛋白,它与TLR亚单位相互作用,形成信号转导复合物。这个复合物激活IRAK家族蛋白,引发NF-κB和MAPK途径的激活。NF-κB调节炎症相关基因的转录,引发炎症反应;而MAPK途径调控细胞功能变化,如细胞增殖、凋亡和细胞因子产生。因此,MyD88在启动下游信号传导和引发炎症反应中扮演着关键角色 [8] [9] 。本研究的目的是通过使用高脂饮食诱导肥胖小鼠模型,研究其脂肪组织中TLR6及MyD88蛋白的表达变化,以探究TLR6及MyD88在肥胖发生过程中是否通过低度炎症的参与起到作用。通过深入理解TLR6和MyD88在肥胖小鼠炎症调控中的作用,我们可以进一步揭示肥胖的发病机制,并为未来的人类肥胖患者治疗策略提供重要的科学依据。
2. 材料与方法
2.1. 实验动物分组及其模型建立
从宁夏医科大学动物实验中心购买4周龄健康雄性C57BL/6J小鼠共计20只,体质量为(14 ± 2) g。将小鼠随机分为正常对照组(NC组,n = 8)和肥胖组(OB组,n = 12)。NC组接受普通饲料喂养,而OB组接受高脂饲料喂养,饲料均来源于江苏省协同医药生物工程有限公司。高脂饲料的配比为60%普通饲料、12%猪油、5%花生、10%鸡蛋、5%奶粉、5%蔗糖、2%食盐和1%麻油。经过12周的饲养后,筛选出肥胖模型小鼠,并剔除其余小鼠。所有小鼠都在宁夏医科大学SPF级环境中饲养,环境条件为温度(22 ± 2)℃,湿度(50 ± 20)%,自由饮水和摄食,12小时昼夜循环。整个实验过程符合宁夏医科大学伦理委员会的规定。
2.2. 脂肪组织样本提取,形态学观察
在高脂肪饮食喂养12周后,采用5%的水合氯醛腹腔麻醉后处死小鼠,迅速取出腹股沟部位的脂肪组织,一部分置于冷冻储存管中经液氮冷冻后−80℃冰箱保存,一部分固定10%的中性甲醛中用于石蜡切片制作,经HE染色后观察脂肪组织形态学变化。
2.3. Western Blot检测细胞蛋白表达
将脂肪组织样品放入事先标记的洁净EP管中,每组选取8份样本进行称重,多余的样品储存在−80℃的冰箱中备用。随后,向每个EP管中加入裂解液(RIPA:PMSF = 100:1),在冰上将组织剪碎后静置于4℃下30分钟。使用超声仪对组织进行超声处理,然后以12,000 rpm的速度在4℃下离心25分钟,取中层液体。通过BCA蛋白定量试剂盒测定各组样品的蛋白浓度,并将浓度调整至2 mg/ml。随后,对样品进行100℃水浴蛋白制样处理,每个孔上样15~20 μl,后进行SDS-PAGE凝胶电泳分离蛋白质。电泳后,将蛋白质转移到0.45 μm的PVDF膜上。接下来,在室温下使用5%脱脂奶粉封闭膜,孵育2小时。随后进行1 × TBST缓冲液洗涤,并加入相应的一抗进行过夜孵育。第二天,膜洗涤后,加入相应的辣根过氧化物酶标记的山羊抗兔或鼠二抗,并在室温下孵育2小时。最后,使用GISt020凝胶图像分析仪对膜进行成像,并使用ImageJ软件分析目的蛋白/GAPDH,目的蛋白/β-actin的吸光度(A值)比值反映目的蛋白的相对表达水平。
2.4. 统计学处理
使用SPSS 26进行统计分析,计量数据以均数 ± 标准差(
)表述。不同处理组检验其结果差异使用t检验,若P < 0.05则具有统计学意义。
3. 结果
3.1. 小鼠腹股沟脂肪组织的形态变化
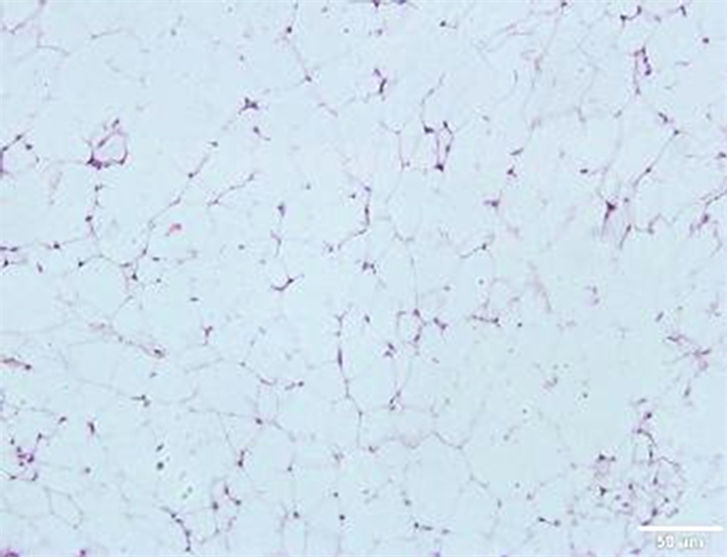
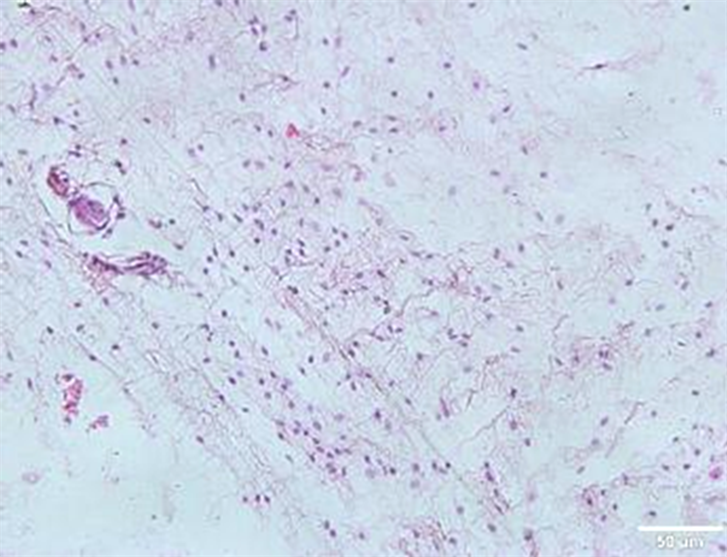
正常组(NC)脂肪细胞形态规整;肥胖组脂肪细胞数量增多,间质内可见多量巨噬细胞见图1。
3.2. TLR-6/MyD88信号通路蛋白在肥胖小鼠脂肪组织的表达
肥胖模型小鼠腹肌沟脂肪组织中TLR6表达水平高于正常组(P < 0.05)见图2;肥胖模型小鼠腹肌沟脂肪组织中MYD88表达水平高于正常组(P < 0.05)见图3。
3.3. IL-6在肥胖小鼠脂肪组织的表达
肥胖模型小鼠腹肌沟脂肪组织中IL-6表达水平显著均高于正常组(P < 0.01)见图4。
 NC组 OB组
NC组 OB组
Figure 1. Morphological changes of mouse groin adipose tissue
图1. 小鼠腹股沟脂肪组织的形态变化

Figure 2. Statistical results of TLR6 signaling pathway protein WB strip gray values
图2. TLR6信号通路蛋白WB条带灰度值统计结果

Figure 3. Statistical results of MyD88 signaling pathway protein WB strip gray values
图3. MyD88信号通路蛋白WB条带灰度值统计结果
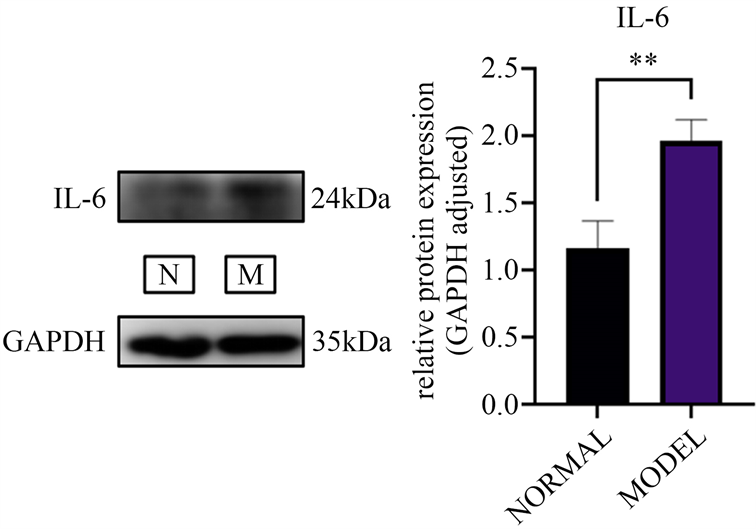
Figure 4. Statistical results of IL-6 WB strip gray values
图4. IL-6 WB条带灰度值统计结果
4. 讨论
肥胖是一种全球性的健康问题,其与许多慢性疾病的发生和发展密切相关。肥胖被定义为体内脂肪积累过多导致身体质量指数(BMI)超过30,并且已经成为心血管疾病、糖尿病、高血压、脂质代谢紊乱等多种代谢性疾病的主要危险因素。此外,肥胖可能导致癌症、睡眠呼吸暂停综合征、骨关节疾病等许多其他健康问题 [10] 。肥胖的发病机制较为复杂,与遗传、环境和生活方式等多种因素有关。在肥胖的发展过程中,脂肪细胞数量和体积的增加是关键因素之一,脂肪细胞的增加可以通过两种途径实现:脂肪细胞分化过程中原有细胞的增殖和新脂肪细胞的形成 [11] 。此外,脂肪细胞代谢活性的改变也会对肥胖的发展起到重要作用,肥胖状态下,脂肪组织中的炎症反应和激素分泌增加,导致脂肪细胞的异常代谢,如胰岛素抵抗、脂肪酸代谢紊乱等 [11] 。
在探讨肥胖发病机制的研究中,TLR6和MyD88等炎症相关因子引起了广泛关注。本研究还发现,在肥胖状态下,脂肪组织中的IL-6水平明显升高,并且在肥胖小鼠腹股沟部位的脂肪组织中发现多量的巨噬细胞浸润,提示在肥胖小鼠的脂肪组织中存在明显的炎症反应,这与文献中报道一致 [12] [13] 。我们还发现肥胖小鼠模型中TLR6的显著表达增加,这可能是由于肥胖状态下脂肪组织中的炎症反应增加,从而引发了TLR6的激活 [14] 。MyD88作为一个重要的信号转导分子,参与调控TLR家族受体介导的炎症反应,TLR6的激活需要与MyD88蛋白结合形成TLR6/MyD88复合物,从而激活下游的信号转导通路,进而促进肥胖的发生 [15] 。值得注意的是,在本研究中,观察到肥胖状态下MyD88的表达水平升高,这可能在这些代谢紊乱和炎症反应中发挥重要作用,进一步加剧了肥胖带来的问题。例如,胰岛素抵抗是肥胖相关代谢紊乱的关键特征之一,在肥胖状态下,脂肪细胞释放过量的脂肪酸,导致胰岛素信号通路的紊乱,使得细胞对胰岛素的反应降低,从而导致胰岛素抵抗的发生。MyD88的升高可能参与了胰岛素抵抗的发展,进一步加剧了肥胖带来的代谢问题。同时,肥胖状态下常伴随炎症反应的加剧,脂肪细胞和免疫细胞释放炎性因子,这些炎性因子进一步激活免疫细胞,如巨噬细胞,释放更多的炎性因子,形成炎症环境。MyD88是免疫和炎症反应的调节蛋白,其升高可能加剧了肥胖相关炎症反应的程度 [16] 。
5. 结论
综上所述,TLR6和MyD88在肥胖发生过程中的作用机制与炎症水平升高密切相关。二者的升高可能通过炎症信号通路促进肥胖的发生。持续的炎症信号激活可导致脂肪组织炎症状态的形成,增加炎症因子IL-6的产生,并进一步参与调控肥胖相关的代谢紊乱 [17] 。此外,炎症反应还可以促进脂肪细胞的分化和脂肪组织的增殖,进一步加剧肥胖的发展。未来的研究可以进一步探索TLR6和MyD88在肥胖病理生理中的具体作用机制,以及炎症信号通路在肥胖发生和发展中的调控机制。这些研究有助于深入了解肥胖的分子机制,并为肥胖的治疗提供新的靶点和策略。
基金项目
2021年大学生创新创业训练计划项目,项目编号:S202110752037。
NOTES
*通讯作者。