1. 引言
鲕粒在碳酸岩中以一种特殊颗粒形式出现。鲕粒内部结构在不同环境中有不同变化,所以鲕粒可作为指向性颗粒使用,对沉积环境的各种变化性能十分敏感 [1] [2] 。原生鲕粒(未受成岩作用改造),其粒度大小和鲕粒含量的多少可以很好地表明温度、生物、流体性质等沉积环境中易变化的因素。若鲕粒经过成岩作用的改造后,鲕粒形态和其内部的构造也能比较清晰地去反映其成岩作用的类型及演化特征。多种不同鲕粒灰岩,尤其是亮晶鲕粒灰岩,这种鲕粒灰岩存在大量的鲕内溶孔是非常有利于油气资源的存储和运移,因此鲕粒灰岩也是一种非常优良的油气存储岩。现代,沉积学学者们对鲕粒在各方面都有着深刻的研究和认识,但在关于鲕粒是如何形成的问题上,仍然存在不同的观点和假说。随着沉积学界的学者对鲕粒的不断深入研究,关于鲕粒成因的认识也愈来愈成熟。国外的研究学者基于对现代鲕粒研究的成果,对鲕粒的成因进行了观点的阐述。国内亦有相关领域的研究学者对鲕粒的成因进行了总结性工作 [3] [4] [5] 。同时,国内拥有十分丰富的鲕粒岩记录,国内学者基于此也在对鲕粒成因的问题进行积极探索和研究。
基于相关鲕粒成因研究的学者研究成果和相关总结的观点,本文将梳理鲕粒成因研究的历史,学习争论激烈的物理化学和微生物的鲕粒成因假说,为今后对鲕粒成因的学习研究提供帮助。
2. 鲕粒成因探索简史
历史上被公认最早对鲕粒进行了系统性研究的学者是古罗马的哲学家、博物学者Pliny the Elder,他将鲕粒描述成一种形似于鱼卵的颗粒 [4] 。17世纪60年代,Hooke对鲕粒进行显微镜下的观察和研究,首次发现鲕粒内部的结构特征,表明鲕粒内部结构是一种类似于鸡蛋拥有的蛋壳、蛋清、蛋黄的同心结构 [6] 。同时,他认为这些鲕粒颗粒的结构是在流体环境中才可能形成的。Brückmann在1721年撰写出版了第一部关于鲕粒岩石的著作,并在其中提出鲕粒是一种被石化的生物结构的观点 [7] 。但是,Da Costa并不赞同这一观点,在多次的研究后,Da Costa指出鲕粒不是起源于被石化的生物化石,而是由于微小的晶体颗粒不间断的结晶形成。同时,反对“鲕粒可以类比鱼籽的形成 [8] ”观点的学者De Saussure表示,在“微小的晶体颗粒不间断的结晶形成鲕粒”的观点的基础上,鲕粒颗粒在流体环境中表面会不断聚集细小的矿物而逐步生长。至此,鲕粒的成因的相关观点出现了生物成因与非生物因素的分歧,这也如一个“小石子”一般,激发了对探索鲕粒成因更进一步的研究与讨论。
随着现代学者对鲕粒的进一步的研究,相关的理论认知逐步完善。学者们对鲕粒是来源于宏体生物的观点提出了批评和否定 [9] 。1823年,De La Beche在牙买加海边的鲕粒进行考察研究时,发现鲕粒需在过饱和碳酸盐的流动水体环境中生长,并表示在鲕粒的形成过程中化学沉淀作用、物理加积作用是很重要的因素 [4] 。研究鲕粒的学者Sorby [10] 在1879年提出“雪球”滚动一样的鲕粒的生长过程的假说,即鲕粒如“滚雪球”一般,在生长过程中可以不断吸附其他微小的矿物颗粒而逐步变大,体现出鲕粒是通过物理作用而形成。由于这种如“滚雪球”一般的假说较困难的解释一些拥有光滑表面(表面光滑较难以将微小颗粒吸附)的鲕粒以及鲕粒纹层呈放射状等系列的问题。从而并未被大多的研究鲕粒的学者们接受 [11] [12] 。同时,在19世纪末,沉积学界的学者已经逐渐关注到“微藻”在鲕粒形成中发挥的作用 [13] [14] 。Wethered [13] 等学者将鲕粒与酸性的溶液反应后发现其中有着蓝细菌残余。Wethered将这些微生物在显微镜下观察时发现他们已经被钙化,并且存在于鲕粒的核心和纹层中 [13] [14] 。基于类似的相关研究,部分学者认为“将鲕粒成因归结为物理作用似乎还为时过早”。 [15] 学者们认为鲕粒成因的探究仍然是个难以解决的问题。2006年Brehn等学者 [16] 耗费4年的时间在实验室里培养出一个“球状微生物群落”,并依此对鲕粒颗粒进行了人工合成,提出了一种以微生物作用形成鲕粒颗粒的假说。经过Duguid [17] 对现代巴哈马鲕粒详细的观察和研究后表明鲕粒在形成的过程中并没有直接的受到微生物作用的影响,同时再次强调了鲕粒的化学成因的理论。生物成因与非生物因素的两种观点在数十年后再次出现争论。
3. 鲕粒物理化学成因的假说
经典的鲕粒物理化学成因假说认为这三个条件为鲕粒的形成所需要的 [18] (图1):
(1) 有核心颗粒的存在;
(2) 碳酸盐过饱和的水体环境;
(3) 一个可以让二氧化碳脱气的动荡的水体 [9] 。
Davies [19] 认为鲕粒是由于无机沉淀形成的,并在不同的水体环境中实验发现,在稳定的环境中形成了放射状的鲕粒,而在动荡的环境中形成了同心鲕粒,表示在动荡的环境中鲕粒表面存在Mg2+离子的附着,阻止了鲕粒表面的生长,而在稳定的环境中鲕粒表面的Mg2+没有附着,从而产生了围绕核心的鲕粒纹层。通过对太平洋部分群岛的鲕粒的研究,Rankey等学者发现现代鲕粒主要形成在高碱度的岛屿附近,在适合的环境下,高pH值有利于使水体的二氧化碳脱气,形成碳酸钙过饱和的条件,促使鲕粒颗粒的发育,从而认为碳酸钙过饱和的条件是限制鲕粒在全球发育和分布的主要因素 [20] 。根据巴哈马鲕粒的研究,Duguid等学者 [17] 发现鲕粒颗粒的形成的过程中并没有直接关联微生物的活动,并通过以下的研究事实来重新强调鲕粒的化学形成的过程。
首先,在巴哈马的鲕粒研究中,鲕粒颗粒都具有极其相似的的化学特征。在鲕粒的内部有较高的氧、碳的同位素含量,而外部的纹层中存在的氧、碳的同位素含量却较低。这种化学特征可以解释为:微生物钻孔中的文石胶结物大部分都存在于鲕粒的内部,从而使鲕粒颗粒内部的碳、氧同位素含量的组成增加 [3] 。再者,大部分的鲕粒外部有较高的Mg/Ca的比值,被称为ACC (Amorphous Calcium Carbonate,非晶质钙碳酸盐)包裹层。同时,外部皮层的Sr/Ca的比值比内部的较低,在化学上造成这种差异的原因是由于鲕粒内部纹层会存在文石胶结物。通过电子显微镜对相应的同位素和化学痕迹元素的分析结果表示,微生物并没有再鲕粒的形成过程中起到相应的关键的作用,相反鲕粒颗粒形成以后微生物的活动却改变了其化学组成和结构。基于此,一种新的鲕粒颗粒的形成模型被建立,当鲕粒处在沉积的阶段时(水面处于活跃,动荡),鲕粒依靠ACC的沉淀薄层开始发育;这个ACC薄层开始结晶形成文石针,再进而形成鲕粒新的纹层(水面处于静止,稳定) [3] 。Duguid等学者 [17] 在综合学者的研究后论述道,鲕粒外部的纹层中氧、碳的同位素含量是不平衡与海水的,可能是由于文石快速沉淀导致的,或是一个文针石的ACC的形成。ACC在很多的生物中会发生生物重结晶,同时,ACC可以重结晶成文石,不仅如此,ACC可以在无机的种子晶体的光学连续中重结晶形成文石 [21] [22] 。在上述的基础上Duguid等学者 [17] 重新提出化学假说,并进一步的阐述其化学构成过程,并将鲕粒的形成过程划分成两个阶段:活跃阶段:鲕粒的皮层开始进行沉淀,表面有较高的Mg/Ca比值ACC;稳定阶段:ACC重结晶成文针石并组成鲕粒的新纹层;Duguid等学者 [17] 认为这种化学过程亦可以很好的解释放射状鲕粒和不规则状鲕粒的形成。鲕粒中大量的微生物痕迹被认为是鲕粒在稳定的阶段让光合作用微生物栖息的场所 [17] 。同时Trower [23] 对鲕粒的生长进行研究发现其生长受到碳酸盐过饱和影响,微生物的微孔化对鲕粒结构起破坏作用 [11] [12] [19] 。
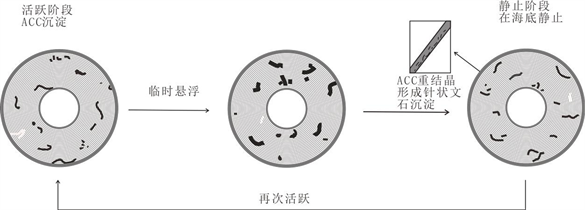
Figure 1. Physical and chemical genetic model of oolites (according to Duguid et al. [17] and Guo Qi Heng et al. [5] )
图1. 鲕粒的物理化学成因模型(据Duguid等学者 [17] 和郭芪恒等 [5] )
基于前文所阐述的内容,鲕粒的物理化学成因的假说可能是周期间断式围绕鲕粒核心的ACC圈层钙化成鲕粒的纹层,形成鲕粒需要高碱度以及有着核心颗粒等不可或缺的条件。鲕粒中含有的大量微生物痕迹被认为是微生物在鲕粒外部繁殖所造成。部分研究学者认为在实验室中人为合成的鲕粒和巴哈马的鲕粒是一个无机的形成过程 [19] ,但这并不是没有有机物的参与,若鲕粒要进一步的发育生长是需要有机膜层提供含能量的基质。Duguid等学者 [17] 重新提出的化学假说被地球化学和SEM调查鲕粒所肯定和支持,其提出的以ACC为模板的形式的前提是鲕粒的皮层最开始是一个ACC的钙化薄层。Duguid等学者 [17] 并没有解释ACC的形成和来源,是他们提出的新化学假说的一个遗憾和理论的缺陷。
4. 鲕粒微生物成因的假说
研究学者在对鲕粒进行微观下的观察研究时,发现地层中发现的鲕粒;正在水体中沉积的鲕粒,都可以在微观下发现微生物的痕迹 [13] [14] 。学者们在不断的研究下逐步产生了鲕粒微生物的假说。
Summons [24] 在1978年便认为特定种类的微生物群落在鲕粒皮层上繁殖有可能促进鲕粒皮层的生长。Brehn等学者 [16] 在学术研究时发现一类不同寻常的球状微生物群落,这种微生物的群落由丝状的蓝细菌以及一种十分特定的方式关联排列的硅藻和异养细菌组合而成。他们在这些群落种发现类似于鲕粒的钙碳酸盐沉淀的作用,并以此提出了一种系统性的鲕粒微生物形成假说 [16] 。历经4年的实验和培育,Brehn等 [16] 培养出了一个微生物的球状群落,并且观察了这个群落的钙碳酸的沉淀,并总结出以下的结论:
(1) 这类微生物群落是由蓝细菌,硅藻以及异样细菌的共生组合。
(2) 这种微型生物球的形成和结构有着相同的规则,其中的异养细菌和硅藻形成一个外部的包裹层,蓝细菌进行靠近并在内部排列,这个球状微生物群落可以存在数月时间。
(3) 在这种微生物的群落的形状是由其钙化作用形成的,在反应中的其聚集颗粒在形成时产生了这个微生物的聚集的群落。在实验中可以观察到同心状的空心球状沉积的碳酸盐晶体的薄层。这种空心层的球也许就是鲕粒的前体,这代表了在没有核心的层状微生物的群落的背景下,产生鲕粒的实验体系 [3] 。
(4) 在球状的微生物的群落进一步的钙化会产生一个稳定是三维的钙化的球体,并在形成完好的球体上反复的发生这种钙化叠加,而在地层岩石中所存在的鲕粒的球状结构就类似这个钙化球的形式 [3] 。
(5) 这种球状的聚合体就是鲕粒,表示出先生成的单个薄层球状结构会在晚期聚合成多层的球状的碳酸盐体。在进行与鲕粒的化石薄片的比对后,可以推断,鲕粒岩石的形成是由于这些球状结构被数次冲刷后再鲕粒滩里大量的聚集 [18] 。
Brehn等 [16] 再实验中没有表明固体核心是微球状的生物群落和鲕粒的形成的必须的条件,但在近代的发现和化石鲕粒的研究中发现鲕粒是将球粒,石英颗粒和生物碎屑作为核心并围绕其所形成的。为了解开鲕粒成因的神秘面纱,科研学者对鲕粒的研究始终没有停下步伐。Tan [25] 发现大量的生物遗迹存在于早寒武系的鲕粒化石中,说明在微生物群落和过饱和的钙碳酸盐的作用下可以形成反射状的鲕粒,表示鲕粒形成中微生物也扮演了重要的角色。Diaz [26] 研究并表明微生物活动可以导致pH和过饱和碳酸钙的局部变化使ACC发育,着重的表示了ACC在鲕粒发育形成中的作用,与熟知的成核经典理论背道相驰,通过ACC是预核化使鲕粒发生矿化,在其中ACC与细胞外聚合物(EPS)和微生物之间的关联十分密切,表示ACC沉淀是由生物介导的,并将鲕粒的形成表述为以下阶段:最初,鲕粒的外部的EPS导致ACC的沉淀形成;ACC在外部累积成薄层;ACC再转变成文石晶体,形成新的文石薄层;外部动荡的水体环境磨损不均匀的文石薄层使其形成光滑的球状,构成鲕粒的纹层。微生物假说有着众多的研究结果支持,众多的学者也都支持这一假说 [3] [5] [27] 。
5. 鲕粒成因观点的讨论
随着鲕粒研究的逐渐深入,鲕粒成因在生物成因和非生物因素两大观点支持基础中逐渐清晰明确。这两种理论首先都要解决ACC的来源问题,并探讨微生物对鲕粒生长所起的特定“角色”。
5.1. 形成鲕粒的ACC的来源的解释
如前所述,在Duguid等学者 [17] 所提出的新化学假说,其中最为重要而又亟待解决的就是ACC自身如何形成。Addadi [21] 等表示:纯净的ACC是处于极度不稳定的状态的,但部分生物可以产生较稳定的ACC (ACC在晶质文石前体过渡中表现出稳定作用)。有关生物ACC研究发现ACC处于稳定状态下含水,过渡形态不含水。与此同时,ACC的结构也存在着显着的不同。现有研究中显示软体动物年幼阶段以ACC为前体,不受外界环境发而发生生物矿化,ACC生物矿化呈稳定状。这表明ACC是生物矿化作用下的一种较特殊的矿物形式以钙碳酸的形式矿化 [28] 。在基础事实上,生物的矿化作用是在新远古代的末期才有所发生的 [29] ;在前寒武至寒武纪,沉积物和沉积面之间存在着一个重要变化被称为“生物骨骼化”(与生物矿化作用和生物扰动相关) [28] 。若将形成鲕粒皮层的ACC薄层表示为含有有机大分子和水分子的矿物,那么Duguid等学者 [17] 表述的鲕粒成因似乎与微生物有一定的相关;被生物矿化的ACC可以形成于生物体外部 [28] ,这一种形成鲕粒皮层ACC的机制与酶的控制无关,同时,这种ACC (含有有机大分子和水分子的钙碳酸盐)亦可以由微生物的活动痕迹产生,这种ACC在蓝细菌的分泌在细胞外的聚合物中尤其常见 [28] [29] [30] [31] 。考虑到Reitner和Arp等学者的研究结果 [30] [31] ,还不可以将Duguid等学者 [17] 重新提出的化学形成的假说理解成鲕粒的形成是一个极其纯粹的化学作用的过程。因为形成鲕粒皮层的ACC可能来源于微生物的作用形成。同时,在现代鲕粒中发现的微生物钻孔也似乎在表明形成鲕粒的ACC皮层与蓝细菌这类微生物的活动有千丝万缕的关系 [3] (图2)。
5.2. 形成鲕粒的ACC的来源的解释
鲕粒颗粒生长过程中,微生物作用可细致归类为破坏性与建设性。存在于鲕粒颗粒内部的非光合作用微生物通过形成代谢等生物活动吸收鲕粒颗粒内部有机质破坏鲕粒内部纹层。与此同时,一些细菌通过生物活动释放出氢离子使得鲕粒颗粒所处环境酸性增加溶解了鲕粒内部碳酸盐,从而产生微小钻孔 [32] 。非光合作用微生物主要表现为对鲕粒颗粒内部原始沉积结构造成强烈侵蚀。微生物建设性作用体现为微生物在其活动中直接参与鲕粒颗粒沉积结构的构建(鲕粒纹层除外)。葛万菌和其余蓝绿藻类等光和作用微生物生长时从鲕粒周围吸收二氧化碳,使其所处的环境碱性增加,另外这些微生物还能分泌细胞外聚合物(EPS)以促进碳酸盐沉淀。
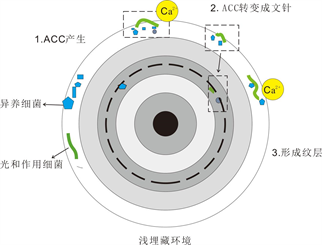
Figure 2. Biogenic ACC model of organic mineralization (according to Li Fei et al. [4] )
图2. 有机矿化生物成因ACC模型(据李飞等 [4] )
6. 建议的鲕粒形成模式
通过对鲕粒成因的学习和更进一步的认识,鲕粒颗粒生成方式可能不是单一物理化学或者微生物生成方式,微生物生命活动能够控制鲕粒外围Ph值(酸碱度)大小以调控碳酸钙析出去影响鲕粒圈层生成,但不同水动力可导致鲕粒颗粒圈层结构变化不一致。郭芪恒 [5] 等人在对前人所提出鲕粒形成依据上将鲕粒形成模式的总结中,对鲕粒的形成作了分类(图3):
(1) 稳定阶段:微生物在鲕粒颗粒生长发挥建设性作用,形成生物群落生成生物膜并分泌EPS (细胞外聚合物);
(2) 活跃阶段:鲕粒颗粒表面有较高的Mg/Ca比值的ACC (Amorphous Calcium Carbonate,非晶质钙碳酸盐)包裹层;
(3) 稳定阶段:ACC重结晶围绕鲕粒的核心针状生长形成鲕粒的圈层结构。若水体的动力环境发生变化会使ACC重结晶围绕鲕粒的核心针状生长的矿物变成切线状或杂乱分布的结构,继续重复(1),(2)进行鲕粒的生长,若水体动力环境未发生明显变化则直接重复(1),(2)进行鲕粒的生长。

Figure 3. Suggests the genetic model of oolitic formation model (according to Guo Qi et al. [5] )
图3. 建议鲕粒形成模式的成因模型(据郭芪恒等 [5] )
在郭芪恒等 [5] 总结的模式表明鲕粒初始圈层应为发射状结构,在生长过程中受到水动力的变化从而导致其他类型的圈层结构的产生。
7. 结论
鲕粒作为碳酸盐岩包覆的重要类型颗粒之一,它的成因问题一直是沉积学界倍受关注的问题和地质学者们关注的焦点。在循序渐进的实验研究中逐渐形成了微生物学说与化学作用学说两种截然相反的看法。但本文经过资料调查与研究,认为鲕粒形成方式不能简单地分为完全化学作用或者微生物作用两种类型可能是共同作用的结果,其形成过程复杂多变。如果要分析鲕粒成因更具体的机制,就需要进一步更加细致地科学研究水体环境控制以及微生物对鲕粒的影响。
基金项目
本文受到宿州学院博士科研启动基金项目(编号:No.2017jb01);安徽省大学生创新创业训练计划项目(S202310379185);宿州学院大学生创新创业训练计划项目(编号:No.KYLXYBXM22-078和KYLXYBXM22-080);宿州学院教师应用能力发展工作站(编号:2020XJYY08);宿州学院“水文地质实验实训中心”(编号:szxy2022sxzx01)的支持。