1. 引言
获得性遗传就是生物后天获得的性状能够遗传给后代。关于获得性遗传虽然存在众多理论与证据,但都不能“盖棺定论”,还存在许多争论。
相关证据有:1、草履虫遗传实验 [1] 。2、枯草杆菌遗传实验。3、蓝绿藻遗传实验。4、高脂饮食诱导形成的跨代传递 [2] [3] 。5、苯乙酮对小鼠的影响 [4] 。相关理论主要是实验中发现的甲基化及RNA对遗传性状的影响。
其它客观现象比如我们对蛇、蜘蛛等等的恐惧 [5] ,一些生物与环境相适应的“遗传性”行为(比如蜜蜂的“跳舞” [6] 等等),如果我们仅仅用基因突变、自然选择来解释,总觉得概率太低,严重缺乏说服力(以人对蛇、蜘蛛先天恐惧为例,人要先天获得这种恐惧的能力,要使大脑中图形信息与恐惧先天建立一定的联系,人的神经细胞必然存在相应的基因及基因功能。而要产生这些基因及功能,首先要产生基因突变。在人的成千上万的基因中,要“刚好”使大脑中图形信息与恐惧成功建立一定联系的对应的一系列基因产生突变,而且这种突变获得的碱基顺序是正确的,大家可以算算其概率。何况,还要从无穷的图形信息中刚好挑出一副所需要的图形信息与恐惧中枢建立记忆联系。最后,产生这些突变的生物还要能够适应环境,并能够将这些突变遗传给后代……。其概率几乎无穷接近0。类似的进化的概率问题,是有些人攻击进化论的理论之一。),但在以上基础上如果再加上获得性遗传,无疑更有说服力。
非生殖细胞繁殖产生后代的生物,产生的获得性遗传现象在理论上相对容易理解。获得性遗传可以分为学习获得的获得性遗传,和“程序反应性”的获得性遗传。气味分子与小鼠那个实验就是典型的通过学习获得的获得性遗传。
1、某些生物在环境刺激下,可以通过核心遗传物质(DNA、组蛋白等等)的改变,产生获得性遗传。比如“蓝绿藻遗传实验”,我们可以解释为环境诱导下,其DNA发生“反应性”变化,从而产生获得性遗传。
2、在环境刺激下,通过相关遗传物质(能够直接或者间接影响DNA的物质称为相关遗传物质。可能有些相关遗传物质,能够在细胞分裂时被大量合成,并按一定规则分配到子代细胞,从而能在一段时间内持续影响遗传)的改变,产生获得性遗传。对于“高脂饮食诱导形成的跨代传递”就可能有这种情况存在,它虽然是生殖细胞繁殖带来的获得性遗传,但高脂饮食通过直接或者间接影响相关遗传物资来影响相关遗传物资。
以上情况下,获得性遗传要长久存在,需要所有获得性遗传都必然有相关和、或者核心遗传物质的改变并影响子代。1) 核心遗传物质的改变带来的情况:只要核心遗传物质的改变,在细胞分裂时能够被复制分配到子代(什么样的改变能够被复制并保存下去?),并能够存在下去,便能够使获得性遗传保持下去。2) 遗传相关物质(比如某些RNA、蛋白质等等)的改变要产生获得性遗传,需要反馈机制:即这些物质要影响核心遗传物质需要保持一定的浓度。由于细胞分裂使其浓度迅速下降,如果不存在相应的反馈机制来维持相对浓度,获得性性状便无法遗传给子代。3) 以上两种情况都具有。(对于高脂饮食那个例子,如果基因活性高低的关键是DNA等的甲基化,那么高脂饮食要产生获得性遗传,要么是DNA等的甲基化能够被复制遗传下去,要么是某些能够直接或者间接影响基因活性的遗传相关物资,能够在细胞分裂的过程中持续保持相对浓度。)
通过生殖细胞繁殖产生后代的生物,对于获得性性状如何改变生殖细胞的基因,并如何遗传给后代,现有的理论是无法给出答案的,需要我们根据现有的实验数据发展相关理论。
一种情况是环境直接带来生殖细胞遗传物质的改变(比如“高脂饮食诱导形成的跨代传递”也可能有这种情况存在)。另一种情况是:环境先带来组织器官中功能细胞的改变,组织器官中特殊的功能细胞再带来生殖细胞的改变,这种情况往往也是学习获得性遗传的情况。第一种情况前面已经讨论,比如高脂饮食诱导形成的跨代传递,存在机理这种可能:高脂饮食会产生某些物质,这些物质既能直接或者间接影响体细胞的基因表达,也能直接或者间接影响生殖细胞的基因表达。(比如实验中发现的RNA等等可能就是这样的物资 [2] [3] 。)
第二种情况是本文重点讨论的问题。“苯乙酮对小鼠的影响”的实验及用进废退(如果存在)便是第二种情况。缺点是这样的实验数据太少。
2. 获得性遗传的进化意义
一种生物一旦进化获得:其通过实践而获得的某些与环境相适应的至关重要的性状,能够遗传给后代的能力。其后代在类似的环境下无疑会更快的获得这些与环境相适应的性状,从而大大提高其后代的生存能力(试想如果某种生物能够通过恐惧遗传,将逃避某种严重危及生命的事件的能力部分遗传给后代,这种生物的后代便不需要反复试错就能够“迅速”学习获得逃避那种危险事件能力,而不具有这种遗传能力的生物,其后代便会在反复试错的过程中大量死亡。在广泛出现这种事件的环境中,这两种生物的生存能力显然会区别巨大)。生存能力的提高,会使一些进化产生的性状更容易的保存下来,而其中一些可能是重要的进化(这些性状开始可能会影响生物的生存),这些重要的进化在未来可能大大的促进生物的进化。因而生物的生存能力的提高会促进其多样性发展,特别是生态系统中的生物刚进化获得获得性遗传能力后的一段时间内,生物的多样性可能会爆发性增长(许多以前不能存在的“变异”生物,因生存能力的提高,而能够存在下去),从而加速生物的进化。
上面的论述可以这样解释:一种生物,当没有获得性遗传的能力时,它具有A群性状能够生存,而当它的DNA因某些原因发生改变而获得B群性状中的某些性状时,它便很难生存下去。但当它因获得获得性遗传而使生存能力增强后,再获得B群性状中的某些性状时,便能够生存下去。这一类生物进化产生B群性状后,便可能进一步进化产生B1、B2、B3……,在这些进化稳定下来之前,便可能使生物的多样性产生爆发性增长。
总之,生物一旦进化获得获得性遗传的能力后,生物的进化便相对是“轻舟已过万重山”。
3. 获得性遗传需要的细胞的结构功能
下面让我们来讨论通过生殖细胞繁殖产生后代的生物,如果要产生获得性遗传的能力,可能需要什么样的细胞结构与功能,同时这样的细胞结构与功能通过进化产生的概率究竟有多大(如无特殊说明,下面的讨论都是针对:通过生殖细胞繁殖产生后代的生物的获得性遗传问题)。
由于对细胞核内物质的运动分布的研究资料不多,我这里只有根据现有的资料,针对细胞的结构与功能做出一些最有可能进化产生的推理与假设。
3.1. 获得性遗传需要什么通路
表观遗传告诉我们,某一性状的改变是相应细胞的相应功能改变引起的,而性状的遗传必然有相应基因活性的改变 [7] 。
就“苯乙酮对小鼠的影响”这个实验来说,这个实验显示,气味分子能够刺激小鼠相应感觉通路并与恐惧对应的神经内分泌建立记忆联系,而且这些对应联系的增强都能够遗传给下一代。在气味分子与恐惧建立记忆联系的神经通路中,刺激联系会使相应突触产生短期记忆、长期记忆,从而使特殊气味分子的刺激与恐惧建立记忆联系,而这些记忆联系要成为终身记忆,就需要这些相应突触后膜的神经细胞(N)的基因活性(M)发生改变,并能够记忆下来。而其后代也要先天具有这种记忆(神经细胞N),必然是通过生殖细胞对应的基因(M)将这种记忆遗传下来。(也有理论假设少量的气味分子进入血液或者RNA穿梭来影响生殖细胞,但仔细分析,这些可能性都比较小。)
要产生这样的遗传效果,必须要精准的调控相关基因。比如假设a,b等基因活性的增强使a1气味分子对生物的恐惧诱发能力持续存在,而c,d等基因的活性增加使c1气味分子对生物高兴的诱发能力持续存在。这个实验中,小鼠通过恐惧记忆,使气味分子能够通过改变相应神经通路中的相应神经的相应基因(是a,b等基因,而不是c,d等基因)的活性来改变精子细胞相应基因(a,b等基因)的活性来实现获得性遗传。而不是像高脂饮食那样,高脂饮食可能直接“先天”改变精子细胞的生存环境,来改变基因的活性,而生殖细胞中a,b等基因活性增加的环境显然无法由气味分子直接“先天”诱发产生(否则也就不需要恐惧学习记忆了)。
小鼠要具有这种遗传能力,需要这个通路中的某些特殊细胞能够分泌某些特异的物质来直接或者间接影响生殖细胞中特定基因的活性。而要从成千上万的基因中精确影响某几个目标基因而不是其它的基因,必须这些物质要携带这些恐惧相关记忆对应基因的“结构标志信息”(只对相关基因进行调控)、大概或者精确空间位置的标志信息(要找到这些基因)。分泌的物质要携带这些信息特异的与遗传物质结合,它应该是具有一定分子量的复杂物资,蛋白质是最有可能的选择。
3.2. 获得性遗传的可能通路
描述这个最有可能的过程:生物的某一功能细胞的某一基因的活性改变的同时,会在这个细胞或者相关细胞合成一种mRNA,它与其结合的出细胞核物质携带这一基因的特异标志信息,在它及其结合的相关物质共同影响下,翻译编辑合成的蛋白质也携带标志基因的特异信息(基因的结构标志信息或者与这个基因的功能特异性相关的结构的标志信息,及基因的空间位置信息——基因在细胞核内的特定染色质区域的某个精确或者大概位置)及其它信息标志(比如可能有生殖细胞所在组织信息或者生殖细胞的标志信息等等)。分泌出细胞后,它包含的信息标志使其能够大量进入循环系统,通过循环系统特异的分布到生殖细胞所在的组织,并与生殖细胞特异的结合,随后有两种途径,一是直接被胞饮入生殖细胞,运输到细胞核,进入细胞核,富集分布到这一基因所在的空间,与这一基因特异性结合。另一途径是这一物质诱导生殖细胞产生相应细胞内物质,它携带这一基因的特异标志信息,并被运输富集分布到……。上述大概过程如图1:1代表细胞核。2代表染色质区域。3代表合成与编辑蛋白质的细胞器。4代表的是分泌出的基因活性调节蛋白。
实心箭头表示:功能组织的相应特殊细胞的功能相关基因参与形成的mRNA复合体被运输出细胞核后,先放射状运输到合成与编辑蛋白质的细胞器,然后编辑成熟的基因活性调节蛋白被放射状运输分泌出细胞。出细胞的基因活性调节蛋白运动分布到生殖细胞细胞膜对应部位,入细胞后(或者是细胞膜上脱离的携带相应信息的分子)被放射状运输到功能相关基因所在的染色质区域对应的部位,从而调节对应基因的活性,而且这种活性的改变能够遗传给下一代。
基因活性调节因子对相应功能基因的调节在神经细胞中可能更容易理解(虽然可以不似对生殖细胞那样的调节)。神经细胞的突触兴奋后,突触前膜分泌的物资带来突触后神经细胞的改变(包括对应基因活性的改变),从而带来短期、中长期记忆的改变 [8] 。这种改变就是类似放射状的(以细胞核为中心,树突为枝干的类放射状)。
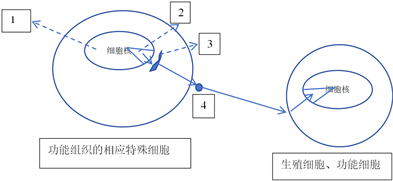
Figure 1. The synthesis secretion of gene activity regulatory factors. And the schematic diagram of its pathway regulating the activity of corresponding genes in germ cells
图1. 基因活性调节因子合成、分泌。以及它调节生殖细胞对应基因活性的路径示意图
功能组织器官中,有多种类型的细胞分泌产生基因活性调节因子,有一种是组织器官中的功能细胞产生基因活性调节因子,另一种是组织器官中调节功能细胞功能的细胞分泌这种物质,还有一种是其它特殊的细胞产生这种基因活性调节因子。分泌调节生殖细胞中对应基因活性的基因活性调节因子的细胞,它的这个功能应该受到这个组织器官中某些细胞的调节。
但不管是那个类型的细胞分泌,翻译合成编辑成熟的基因活性调节因子,必然包含前述的一系列的标志信息。
3.3. 可能通路的一些特点的进一步讨论
3.3.1. 特异性结合的进化产生
如何理解基因活性调节蛋白与对应基因或者基因相关结构的特异性结合的进化产生?我们可以将基因及其相关结构看作受体,而基因活性调节蛋白看作配体,这样既然在原核生物中就存在受体与配体,进化产生类似受体与配体能够特异结合的基因活性调节蛋白与对应基因也是容易理解的。
更进一步,既然在生物体内,抗原能够诱导某些细胞产生抗体,那么mRNA复合体在某些细胞中也可能“诱导”出“抗体”,从而使这个“抗体”与对应基因(这个mRNA对应)能够发生特异性结合,这个“抗体”如果是这个基因的基因活性调节蛋白,它与这个基因或者相关结构特异性结合后,就会调节这个基因的活性。
因而生物进化产生能够与对应基因或者相关结构特异性结合的基因活性调节蛋白的可能性是非常大的。
当然,如果细胞核内的空间结构因子对空间的定位能够精确到基因,在没有本节所述的特异性结合机制的情况下,也可能进化产生这种特异性结合效果。
3.3.2. 生物生存资源对特异性分布的影响
生物体内,如果这些被分泌的物质(基因活性调节蛋白)没有特异性分布,那么,在各个器官组织中可能会相对均匀分布,在目标基因活性被诱导需要的物质(基因活性调节蛋白)浓度不变的情况下,非特异性分布消耗的资源是特异性分布的几倍甚至数百倍。而生物体内参与各种获得性性状的基因数量应该不少,再加上蛋白质的代谢消耗,使分泌的基因活性调节蛋白不具有特异性分布能力的生命如果要获得获得性遗传的能力,消耗的资源无疑会成几何级的增加,使它对恶劣生存环境的适应能力大大下降。
生物体内分泌的某一物资,根据其在生物体内特异分布的特点,可以分为:1、只能在全身分布,无特异性。2、可以特异分布到器官,在器官内的分布无特异性。3、可在器官内某一局部组织内能够特异性分布。在这个器官的这个局部组织内的分布浓度远高于这个器官其它部位的分布浓度。4、能够特异性的分布到这个组织的某一类细胞上(比如生殖细胞)。5、特异性分布到生殖细胞的基因对应的染色质空间。6、特异性分布到染色质空间包含的基因分布的某一空间。
对比全身、器官、器官局部组织、器官局部组织的局部细胞、特定细胞的特点染色质空间、基因分布的狭小空间,所占有的体积可知,特异性分布每精确一步,其特异性分布的空间体积就会呈几何级数的下降,需要的资源(能量、“材料”)更是会呈几何级数的下降。
消耗的资源:……→特异性分布到组织→特异性分布到生殖细胞→特异性分布到基因对应的染色质空间→染色质空间的局部空间→基因分布的狭小空间。
生命在进化的过程中,经常处于资源缺乏下,在资源缺乏下,资源利用效率的提高必然会大大提高生命对环境的适应能力。生物分泌基因活性调节蛋白如果消耗的资源成倍减少,其在恶劣环境中获得有助于生存的获得性遗传的能力就会大大增强,从而使生存能力成倍增加。
在生物生存的环境资源下,生物的某一获得性性状要遗传给下一代,与这一性状对应基因的基因活性调节蛋白的分泌能力、这一调节蛋白对对应基因的调节能力及特异性分布的特点密切相关。如果资源充足、调节能力超强,可能只需要能够特异性分布到器官就可以使目标基因局部的基因活性调节蛋白的浓度达到调节要求。当然如果资源越充足,特异性分布越精确,就会有越多的基因能够产生获得性遗传。
生存适应要求生物相关功能细胞分泌的基因活性调节蛋白的特异性分布越精确越好。
现在的问题是特异性分布能否进化产生。从理论上在各个组织的特异性性分布(指一种物资在某一特定组织的分布浓度远大于其它组织的分布浓度)是能够实现的,一种实现的途径是通过组织特异性抗原(只要一物资能够与某个组织的组织特异性抗原特异性结合,便能在这个组织中富集)。更进一步的特异性分布,下文继续讨论。
3.3.3. 细胞结构的要求
神经细胞因其相互联系的特点及兴奋记忆的差异性(特异性),成为我们讨论获得性遗传一个很好的参考对象。
神经细胞的突触兴奋后,会带来突触后神经元的改变,由于蛋白质及RNA会不断被降解,难以维持突触的更长期记忆,因而相应基因活性的改变是更长期记忆的基础(有试验证明记忆与基因活性的改变密切相关 [8] )。相应基因活性的改变,应该是与局部突触对应的相关基因,而不是所有基因的改变。因为如果一个事件a引起一个神经元的一个突触a1兴奋,而事件b会引起这个神经元的另一个突触b1兴奋,并产生短期及长期记忆。长期记忆的基础是对应基因活性的改变(要记忆不被遗忘,需要突触的相关蛋白维持一定的浓度,而蛋白浓度的维持需要基因转录的mRNA来提供支持)。如果突触兴奋引起的基因活性的改变不具有特异性,那么事件a发生后,就会引起这个神经元所有的相关基因兴奋,从而使b1在没有兴奋的情况下就产生了长期记忆,事件b还没有发生便产生了记忆,显然是不符合现实的。比较符合现实逻辑的通路是:突触兴奋后分子量较高的基因活性调节分子(它可能是被胞饮入的分子,也可能是突触前基因活性调节分子与细胞膜上的分子结合后,而脱离细胞膜的分子)被微管放射状运输到细胞核,入细胞核后“就近”调节相应基因活性,这个基因转录的mRNA“就近”出细胞核,由于微管运输及细胞骨架的阻碍作用,其在放射状区域翻译、编辑、合成相应蛋白质,然后被在放射状区域被运输到这个突触,从而维持这个突触的被兴奋能力。这样能够相对保持突触兴奋后对基因活性调节的特异性。由于微管及细胞骨架系统在细胞中的广泛存在,我们有理由相信,从细胞膜到细胞核的这种放射对应性调控可能普遍存在。
(细胞的一个染色质在细胞核内的分布区域叫染色质域,不同的染色质有不同的染色质域。)
这种放射对应性非常有利于获得性遗传的进化获得。我们可以这样假设:由于微管的运输作用及细胞骨架系统对细胞内物资特别是大分子物资的运动限制作用,一染色质区域某一空间局部(a)与细胞膜的某一局部区域(b)存在放射状联系,如果这一放射状区域合成的某些蛋白质携带标志a的信息,那么进化可能使:a区域基因产生的携带标志a信息的基因活性调节蛋白分泌后,可能与其它细胞的细胞膜的b区的携带有标志a信息的蛋白质作用,然后携带标志a信息的物资(胞饮进入或者从细胞膜上脱离形成)被运输到这个细胞细胞核的a区,从而完成对相应基因的调控。
3.3.4. 空间分布的特异性,空间因子
下面讨论的是在前文基础上,更进一步的特异性分布的可能性。
1) 基因特异性空间分布的可能性
我们知道细胞核位于细胞中央,核仁位于细胞核的中央,各种细胞器都有其大概的空间位置……。我们可以从使这些细胞内结构,特异性分布到相应空间的因素中抽象出标志它们空间位置的信息。有的物资会特异性分布在细胞核内,有的物资会特异性的分布在相应细胞器内。与染色体对应染色质有特异性空间分布吗?更进一步,基因能够有其特异性的分布空间吗?
细胞核中在染色体固定到细胞核膜上后,异染色质解螺旋成为常染色质的过程中,常染色质链在细胞核中是如何分布的?杂乱分布?这样会不会发生染色质的不同部位纠缠在一起而影响到染色质的翻译、复制、分离,而影响细胞的一系列功能。如果染色质是有组织的空间分布,染色质链应该携带空间标志,使不同的染色质链有组织的分布在细胞核相应空间中,这样即使染色质链因为各种原因发生断裂,也很容易修复(由于空间标志没有改变及细胞核骨架的影响,因而断裂而分开的两部分不会发生远离,容易在适当的情况下重新连接,并使错误连接的可能性大大减小),在复制后螺旋压缩形成染色体的过程中,即使有纠缠断裂,染色体各部分在空间标志影响下也容易“找到自己的空间位置”。那些没有空间标志的染色体片段相对于那些有空间标志的染色体片段,在解螺旋,复制,细胞分裂的过程中更容易丢失,从而使染色质片段一般都有其空间标志。
如果DNA片段有空间标志,那么DNA片段的空间标志在哪里?是垃圾DNA还是……,等等结构。这些空间标志与空间因子(或者其本身就参与空间因子的组成,空间因子可能是蛋白质或者各种RNA)结合,从而影响DNA片段在细胞核内的分布。
我下面设想一种可能的情况:在细胞核中是否存在一个空间因子集合,每个空间因子可以对应DNA特殊位置上的一组或者一个碱基,空间因子之间及空间因子与环境之间能够相互作用而决定空间因子相互的空间位置,一群空间因子可以根据DNA某一片段上特殊部位的一群碱基按照一定规律组合在一起形成影响DNA片段空间位置的聚合体,不同DNA片段对应的DNA空间聚合体相互作用,而决定了染色质上一系列DNA片段的空间位置,这样的空间因子及DNA空间聚合体能够进化产生么。如果进化产生了,那么每个染色质(染色体)在细胞核中都会有它的固定空间,而染色质上的每个DNA片段在这个染色质空间都会有其相对固定的空间位置。
2) 空间标志信息的转换
如果在细胞核内的每个DNA结构都包含或者结合了空间位置结构因子,这些空间位置结构因子相互作用,使不同的DNA片段都有其特异的相对固定的空间位置,当然也存在特异的空间标志(a)。在基因合成mRNA后,其出细胞核的mRNA复合体,被按照一定规则赋予了与这个空间标志(a)相对应的空间标志(这个空间标志可能精确,也可能粗糙,粗糙到可能只是染色质空间的标志,而精确,可能精确到基因所在的狭窄空间),然后这个空间标志又被按照一定规则赋予给了被翻译、编辑、合成的基因活性调节蛋白。合成的携带众多信息标志的基因活性调节蛋白,特异性结合目标细胞细胞膜上相应的蛋白质。
基因活性调节蛋白通过胞吞来实现进入目标细胞的细胞核进行基因调控是效率是比较高的特异性调控方式,而如果通过细胞膜上的蛋白质反应来间接完成特异性调控大概有两种方式:1、基因活性调节蛋白携带的各种信息,需要重新转换,但实现信息转换的结构在哪里?一方面效率低,另一方面会大大提高进化的难度。2、大量能够参与获得性遗传的基因另外合成携带相关信息(特别是能够与一基因发生特异性结合的“抗体”信息)的相应蛋白质分布在细胞膜上,这会消耗大量的资源。这样的蛋白质与“基因活性调节蛋白”结合后,其相关结构(携带相关信息)会脱离那个蛋白质,并被运输到相应位置调节基因的功能。这条通路,基因活性调节蛋白不直接调节目标基因的活性。
3.3.5. 获得可能性性状更容易通过精子细胞遗传给后代
现在的问题是,因为生物不同基因的活性被调节难度可能不同(有的需要更高的基因活性调节因子的浓度)及基因分布的空间位置不同(有的靠近核孔,基因活性调节蛋白可能更容易到达,从而能够有更高的浓度),使基因活性被调控的难度千差万别,而且功能组织中特殊细胞分泌的基因活性调节蛋白的量也可能千差万别。因而对于一个生殖细胞来说,应该只有部分基因能够参与获得性遗传的形成。但对于“无数”的精子细胞来说,有的基因在某些精子细胞中其不容易调控(其活性需要更高的调节因子浓度;更远离核孔,或者其它原因使基因活性调节因子不容易达到其所在空间。),而在另一些精子细胞中,它可能更容易被调控(其活性可能需要更低的调节因子浓度;更靠近核孔,或者其它原因使基因活性调节因子可能更容易达到其所在空间。)。而如果精子细胞被基因活性调节因子调控到一定程度后,使它相对于其它精子细胞更容易被选择存在下来,那么获得性性状应该更容易通过精子细胞遗传给后代(因为精子细胞太多,从而使能够产生获得性遗传的基因更多,而卵子细胞太少,而使能够产生获得性遗传的基因数目相对比较少)。(生物如果进化产生了这种选择精子细胞存在的模式,便能够将更多至关重要的获得性性状遗传给后代,更有利于生物的生存,从而使这种模式被选择下来。)
3.3.6. 组织中可能的特殊细胞
生物进化过程中,一组织中可能会产生一种特殊的细胞,当这个组织的获得性性状对应的基因活性增加时,这类细胞中与获得性性状对应的基因的活性也增加,它转录出mRNA,mRNA复合体携带基因的空间标志,出细胞核翻译编辑成成熟的蛋白质,这类蛋白质携带对应基因的一系列空间标志(所在染色质区域的标志,或者更进一步染色质区域某一基本空间的标志),并能够在分泌后,到目标细胞直接或者间接与这个基因像抗体与抗原(或者受体与配体)结合那样特异性结合。
前面论述了神经组织的突触前细胞能够分泌基因活性调节蛋白,特异性的调节突触后神经细胞对应基因的活性。我们可以这样认识:组织中相关特殊细胞与生殖细胞的关系类似突触前及突触后神经细胞,只是它们没有轴突将它们联系起来,它们的长距离的联系需要给这类特殊细胞分泌基因活性调节蛋白剪接加上相关信息标志,使其能够在生殖细胞所在空间能够富集。通过前文的讨论可知,生物进化产生这样的组织特殊细胞是可以理解的。
4. 什么情况下产生获得获得性遗传,奖惩、应急反应
只有那些有利于生物生存的获得性性状遗传给后代,才能够提高生物生存的能力。因而通过进化,生命可以获得这种能力:一个性状有利于生物生存达到一定程度,生物便能够通过一定规则将这种性状遗传给后代。如果我们把获得的有利于生物生存的状态(环境状态及对应的神经内分泌状态)称为奖赏状态,而把获得的不利于生物生存的状态称为惩罚状态。根据已有的资料可知,一般情况下有利于生物生存的状态,细胞所处的内环境相对适宜,生物代谢旺盛,而不利于生存的状态,细胞所处的内环境相对不适宜,生物代谢下降。(生物进化获得奖赏状态及强烈应急状态下分泌物质调节细胞基因的活性的能力,在我们的神经系统中对应的突触前后的神经细胞中我们就能够看到,突触兴奋后突触前细胞分泌的物资对突触后细胞基因活性的影响 [8] 。)
生物在奖赏状态或者恐惧状态下,其对应的功能组织中活性增加的一些功能基因,在生殖细胞中与它对应的基因的活性也可能被调高。这些功能基因的活性增加有助于奖赏状态的获得,同时这些基因对应的功能活动的增加有助于奖赏状态(逃避惩罚也是获得奖赏)的获得。比如气味分子与小鼠的那个例子,相应功能细胞的相应基因的活性的增加有助于气味与恐惧的长期记忆的建立,从而有助于小鼠逃避相应惩罚。
我们可以这样假设,在生物处于奖赏状态或者恐惧状态下,会分泌一些物质,使那些活跃组织中,与相应活跃功能对应的活跃到一定程度的基因的活性会被调高,并被强化(基因活性的持续增强),而这个组织中相应的特殊细胞,诱导生殖细胞中对应基因活性的能力,也会被强化。
在生物的实践过程中,有些事件需要成为短期记忆,有些需要成为长期记忆,而有些严重的影响到生物的生存,需要改变相应基因的活性,从而成为更长期的记忆,更长期的记忆如果能够遗传给下一代,就能够增加种群生存下去的概率。因而,通过生存选择,那些能够严重影响生物生存的获得性性状,就可能遗传给后代。
我们知道,我们能够适应环境,是因为在环境下,我们机体的众多功能的活性在一个适当的范围内,从而使众多功能相对平衡,这种平衡使我们更能适应环境,也会使我们感觉相对舒适(奖赏状态)。奖赏状态下,各个功能活性状态会得到强化,强化到一定程度会通过获得性遗传(如果我们有这种能力),遗传给下一代,从而使下一代的各个功能活性能够迅速达到相对平衡,从而更好的适应环境。生物进化过程中,通过随机突变产生有益基因的概率是非常低的,有益基因产生的同时要不影响其它基因对应的功能平衡,其概率更低,即使“撞大运”,各个功能能够在活性上平衡,但许多突变获得的功能要正常发挥作用,还要通过学习获得,这进一步降低了生物被选择存在的概率。而获得性遗传能够将至关重要的,通过学习获得的“适当”的功能(这一功能才开始产生时并不“适当”)遗传给下一代。获得性遗传为生物功能的复杂化准备了条件。获得性遗传能够让产生有益突变的生物更容易获得生存上的优势,从而加速生物的进化。
我们日常可以有意识的学习,对环境的适应仍然不是最优的。对于学习效率远低于有意识学习的获得性遗传的学习,在环境下学习适应的能力应该是非常有限的。这种遗传学习只能是针对严重影响到生存或者反复获得奖赏事件的遗传记忆学习,当然这种学习是非常模糊的。越精确的学习,越会对生物的遗传系统提出更多更苛刻的要求,也需要更多的资源,同时也更难进化产生。
针对获得性遗传更进一步的假设
假设生物的某些基因(a、b等等)的活性能够影响各种蛋白质的剪接模式,在这些基因(a、b等等)相应活性的影响下,会有各种剪接模式,“正确”的剪接模式,会获得需要的蛋白质,有利于获得“奖赏”,从而通过获得性遗传被强化。这就像生物的行为模式,通过学习记忆获得的行为模式,更丰富更有弹性,更能适应多变的环境,且更容易获得,而通过进化固定的行为模式,都比较简单而没有弹性,更难获得,对环境的适应能力也差了许多。我们可以想象,在进化获得某些功能基因的早期,如果能够相对“随机”剪接组合蛋白质,并通过获得性遗传将“正确”的剪接组合模式固定下来,无疑会大大提高生物的生存进化能力。所以当这种剪接学习与获得性遗传建立联系后,生物的进化会大幅度加速。
如果这种机制存在,对我们理解,高等生物进化产生成千上万的各种功能,而且各种代谢功能、反应功能、运动功能等等都处在相对“合理”(“适当”)的范围内的概率问题能够提供极大的帮助。试想生物基因突变产生一种新功能,这种新功能刚好与其它功能相适应的概率是比较低的,但如果有获得性遗传及这种剪接“奖赏”模式,生物基因突变形成的新功能通过遗传学习便有可能在有限代际繁殖后,迅速与其它功能相适应,并通过剪接获得的延伸功能融入生物的其它功能中,共同促进生物对环境的适应能力。
5. 小结
生物要适应环境,就需要相应的功能,而生物对相应功能的获得可以通过:各种突变产生相应新功能;获得性遗传获得相应新功能(如果本文讨论的获得性遗传规则存在);有意识地学习获得相应新功能。获得性遗传既可能使突变产生的新功能在有限繁殖代际内迅速与环境相适应,也可能使有意识学习获得的至关重要的功能遗传给后代,从而大大提高生物的生存能力及进化速度。
生物要具有获得性遗传的能力,就需要它的某些细胞具有一些特殊的结构与功能。这些特殊的结构功能,主要列举如下:首先什么样的DNA三维结构能够被复制到子代,并能够保持功能活性。或者是什么样的核心遗传相关物资,它们既能诱导基因的活性,又能在子代保证其浓度的相对稳定;细胞核内与染色体对应的不同染色质是否有以核膜为附着点的相对固定的区域。是否有标志染色质区域的信息。细胞核内是否有空间结构因子及相关标志信息,如果有会精确到什么程度。出细胞核的部分mRNA是否遵循就近的原则,翻译编辑合成的部分蛋白质是否始终在放射状区域附近运动分布。以染色质区域为起点,以微管为轴,以细胞膜为终点的放射状区域内的某些蛋白质是否携带这一放射状区域的空间标志信息。细胞膜上的某些蛋白质是否携带这个放射状区域的标志信息。分泌的某些蛋白质是否能够携带特定组织细胞的定位信息。本文所述的精子细胞的选择机理是否存在。……。
研究这些问题除了对获得性遗传有帮助,还对基因工程、肿瘤治疗等等多个研究领域有巨大的应用价值。如果我上面提出的细胞的结构功能都存在,那么我们借鉴细胞的这些功能,通过生物工程获得的药物就可能精确的定位特定组织、器官、细胞、基因,就能够高效、精确灭杀肿瘤细胞,能够高效、精确沉默某一特定基因。……。
获得性遗传对我们理解神经系统的一些学习现象也有巨大的帮助:比如,如果高等生物的a神经集合兴奋b神经集合后一至关重要的行为模式产生,获得性遗传可能会使其后代的a更容易易化兴奋b (比如本文的气味与老鼠的恐惧学习例子),如果这种情况存在,我们就能够容易的解释下面的一些学习现象。1、虽然我们可以通过学习记忆来解释镜像神经元的镜像现象,但如果再结合获得性遗传来解释镜像神经元的镜像现象,将会更加圆满(比如对刚出生的婴儿伸舌头,他的镜像神经元的先天反应)。2、如果有了获得性遗传,婴儿对各种思想行为模式的学习将更加顺利(比如模仿的学习,在婴儿看到某一个行为时,如果与这个行为相关的神经中枢先天更容易被易化,那么必然会大大提高模仿学习的效率)。……。
如果进化没有实现这样获得获得性遗传(本文讨论的获得性遗传)的方式,说明人类还有很大的进化空间,我们未来是否可以通过生物工程改造我们的生殖细胞,来使我们获得这样的获得性遗传的能力。不过根据现有的资料,我倾向于人类有这样的获得性遗传的能力。