1. 引言
植物抗寒的开始是因为环境因素改变了植物体内激素间的相互平衡,导致植株体内各种代谢途径的变化和生长的停止。植物抗寒锻炼的主要推动力是植内源物激素平衡关系的改变。有研究指出[1] ,抗寒基因表达的启动因素就是植物激素,对植物抗寒力重要的调控作用。
赤霉素(GA)是一个较大的萜类化合物家族,在植物整个生命循环过程中起着重要的调控作用。虽然只有部分赤霉素具有生物学活性,但这些有活性的GA不仅与植物的抗寒性有关[2] ,而且调控能够调控植物的各个生长发育阶段[3] 。GA3是最早被认为与抗寒性有关的植物激素。一般认为抗寒性强的植物GA3含量低于抗寒性弱的植物。因为GA3含量下降能抑制生长,提高抗寒性[4] 。GA和IAA为促生长类激素,对植物的生长发育主要起促进作用,逆境条件下植物一般需要降低生长速度以度过不良环境[5] 。
植物内源激素对植物抗寒性的影响方面国内外研究也比较多。植物激素如脱落酸(ABA)和赤霉素(GA3)的调控作用已被许多研究者公认[6] 。多年来的研究,植物抗寒力诱导过程中内源激素的水平使用外源激素来提高,从而能提高植物的抗寒力已被有所证实[7] 。适宜浓度的GA3可以降低O2−生成速率、MDA含量,增加脯氨酸、可溶性蛋白质含量,延缓芍药的衰老[8] ,提高黄瓜幼苗叶片POD、CAT、SOD的活性,降低质膜透性,缓解了黄瓜幼苗的盐害[9] 等。外源GA3也可以显著降低槭树的抗寒力[10] ,上述结论在冬小麦[11] 、苜蓿[12] 、柑橘[13] 等植物上均得到证实。
近年来在果树生产上使用外源激素来提高果实品质、生长发育、开花结果、育种、果实无子化等研究成果也比较多。也可以看到外源激素减少花期冻害的研究报告。目前外源激素应用在果树研究比较多的是草莓[14] 、樱桃[15] 、核桃[16] 、苹果[17] -[19] 、红枣[20] [21] 等果树的生理功能、品质和提高繁殖力的影响方面,杏[22] -[24] 在抗寒性关系研究,葡萄糖代谢、果实品质提高[25] -[27] 等方面有报到。
库尔勒市自从进入90年代以来,冬季发生了多次大范围的冻害,基本是3~4年一遇,冻害已成为香梨产业健康发展的第一大障碍。本研究针对以上生产上急需问题,提高库尔勒香梨抗寒性为目的,在展叶期对香梨树叶面喷施不同浓度不同次数的外源赤霉素(GA3)。并在休眠期枝条取样,结合人工低温胁迫条件下测定其有关抗寒指标,探讨展叶期外施赤霉素对库尔勒香梨抗寒性的影响。旨在为库尔勒地区的香梨安全越冬、稳产和高产提供理论依据。
2. 材料与方法
2.1. 材料及前期处理
本试验在库尔勒市阿瓦提乡进行。选择管理较好的15年生的库尔勒香梨果园中树冠大小和树势相对一致的30棵香梨树确定为试验材料。本试验所用外源赤霉素(GA3)是美国Sanland Chemical Co, LTD生产,纯度大于96%。
2013年5月6日第一次叶面喷施赤霉素(GA3),处理浓度为:50 mg/L、75 mg/L、100 mg/L,记为A1、A2、A3;隔20天后第二次喷施相同浓度的GA3,记为B1、B2、B3;隔20天后第三次喷施,记为C1、C2、C3。共9个处理,每个处理各选取3棵树重复试验,共27棵树;对照处理喷施清水,记为CK,同样选取3棵树重复。其他管理条件一致。
2.2. 试验处理
2013年12月30日取样,各样品均选取树冠外围生长健壮、粗细均匀一致的一年生枝条。取回的一年生枝条先用清水洗净,再用蒸馏水冲洗。擦拭晾干后,按照5个人工低温处理温度,将每个处理枝条平均分成5组进行实验。用洁净湿纱布包裹试样,放入低温冷冻箱进行人工低温胁迫处理。处理温度为:−20℃,−22℃,−24℃,−26℃,对照温度为4℃,每个处理冷冻12 h以上。冷冻结束后,在4℃下解冻30 min,然后将枝条用于测定抗寒有关生理指标。
2.3. 指标测定
膜透性采用电导法测定,丙二醛(MDA)含量的测定采用硫代巴比妥酸(TBA)法测定,游离脯氨酸含量测定采用酸性茚三酮染色法,可溶性蛋白质含量的测定采用考马斯亮蓝G-250染色法,可溶性糖和淀粉的含量测定采用蒽酮法。
2.4. 抗寒性评价
抗寒性综合评价用隶属函数法综合各项指标进行抗寒性评价[28] ,其公式为:
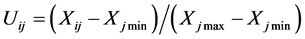 (正相关)
(正相关)
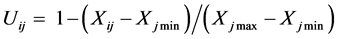 (负相关)
(负相关)
式中:Uij表示i种类j指标的抗寒隶属函数值;Xij表示i种类j指标的测定值;Xj min表示所有种类j指标的最小值;Xj max表示所有种类j指标的最大值;i表示某个品种;j表示某项指标。
按照平均隶属度将抗寒性分为5级[29] :0.70~1.00为高抗(High Resistance, HR),定为Ⅰ级;0.60~0.69为抗(Resistance, R),定为Ⅱ级;0.40~0.59为中抗(Mid-dle Resistance, MR),定为Ⅲ级;0.30~0.39为低抗(Lower Resistance, LR),定为Ⅳ级;0~0.29为不抗(Susceptible, S),定为Ⅴ级。
2.5. 数据处理
实验数据分析采用DPS7.05统计分析软件完成。
3. 结果与分析
3.1. 外源GA3处理对各抗寒指标的影响
3.1.1. 外源GA3处理对库尔勒香梨枝条膜透性的影响
外源GA3处理后,人工低温条件下,总体上各处理库尔勒香梨枝条的相对电导率随着温度的降低而增加,但增加的幅度并不均匀。
由表1得出,各处理温度下各处理之间的差异性达到了显著水平。人工低温胁迫下,在降温至−24℃时,除了B2、C2处理外,各处理的相对电导率均小于对照CK。C组处理在−22℃时急剧上升,此后增幅缓慢。人工低温胁迫至−26℃时,除A2处理外,各处理相对电导率都有明显的增加,均比4℃时增加31%以上。C2的增加幅度最大,为56.12%,A3次之,为55.38%。对照上升幅度为24.05%。各处理浓度的电导率在人工低温胁迫下增加的幅度不一,这也说明了低温处理枝条内的膜透性发生了异常变化,各处理的抗寒效果表现均不同。
3.1.2. 外源GA3处理对库尔勒香梨枝条MDA含量的影响
丙二醛(MDA)是膜脂过氧化的最终分解产物,具有细胞毒性,对质膜有毒害作用,主要破坏细胞膜系统。不同浓度外源GA3处理,人工低温胁迫条件下,总体上各处理库尔勒香梨枝条的MDA含量随着温度的降低呈先升高后降低的趋势,且变化的幅度并不均匀(表2)。
从表2中可看出,除了−24℃温度下,各处理温度下各处理之间的差异性达到了显著水平。A组各处理在−24℃时MDA含量达到最高值,均比4℃下上升62%以上,其中A3处理增幅最大,为95.68%,−24℃之后急剧下降。说明在−24℃时,A组各处理的细胞膜脂过氧化程度最高。B组和C组在−22℃达到最高值,均比4℃下上升52%以上,其中C1处理增幅最大,为107.41%,−22℃之后急剧下降,−26℃时,C1处理MDA含量降低了60.51%,说明在−22℃时,B组与C组各处理的细胞膜脂过氧化程度最高。
3.1.3. 外源GA3处理对库尔勒香梨枝条游离脯氨酸含量的影响
各处理温度下各处理之间的脯氨酸含量差异性达到了显著水平,4℃温度下除外(表3)。不同浓度外

Table 1. Effect of branches conductivity from Korla Fragrant Pear under the treatments with different concentrations of Exogenous GA3 (%)
表1. 外源GA3处理对库尔勒香梨枝条电导率的影响(%)
注:同一列数据后小写字母不同者表示差异显著(P < 0.05),下同。
Table 2. Effect of malondialdehyde content from Korla Fragrant Pear branches under the treatments with different concentrations of Exogenous GA3 (µmol/g)
表2. 外源GA3处理对库尔勒香梨枝条丙二醛含量的影响(µmol/g)
Table 3. Effect of proline content from Korla Fragrant Pear branches under the treatments with different concentrations of Exogenous GA3 (ug/g)
表3. 外源GA3处理对库尔勒香梨枝条游离脯氨酸含量的影响(ug/g)
源GA3处理后库尔勒香梨枝条内的游离脯氨酸含量随着低温胁迫的加强有不同程度的增加,−26℃下,A组各处理的脯氨酸含量达到最高值,而B组和C组各处理的脯氨酸含量在−24℃达到峰值后出现下降趋势。在−20℃时,各处理的脯氨酸含量急剧上升,温度继续下降,脯氨酸含量缓慢增加。在低温锻炼下各处理后的脯氨酸含量均比对照低。其中脯氨酸含量最大值出现在−26℃时A3处理,为30.85 μg/g,比4℃处理上升了151.55%。说明同处理条件下A3处理的枝条受到的低温伤害较小。
3.1.4. 外源GA3处理对库尔勒香梨枝条可溶性蛋白质含量的影响
遭受低温胁迫的植物,可溶性蛋白质的含量会有所增加,以此对植物抗寒起调节作用。由表4可看出,随着处理温度的降低,各处理的可溶性蛋白质含量均呈逐渐升高的趋势。且各处理温度下各处理之间的蛋白质含量差异性显著。其中温度在4℃至−20℃间,香梨枝条中的可溶性蛋白质含量迅速增加。−22℃后,各处理枝条的蛋白质含量缓慢增加,在−26℃达到最高值。说明香梨在−22℃开始积累一定的渗透调节物质,以适应外界环境的变化。在−26℃的低温胁迫下,除A2处理外,各处理枝条的可溶性蛋白质含量均比对照高,并且C2处理的枝条的蛋白质含量达到最高值为8.06 mg/g,比同处理中的对照高出21.12%,比4℃的处理上升了319.80%。
3.1.5. 外源GA3处理对库尔勒香梨枝条可溶性糖含量的影响
随着胁迫温度的下降,各处理枝条的可溶性糖含量总体呈先上升后降低的趋势(表5)。−24℃之前各
Table 4. Effect of soluble protein content from Korla Fragrant Pear branches under the treatments with different concentrations of Exogenous GA3 (mg/g)
表4. 外源GA3处理对库尔勒香梨枝条可溶性蛋白质含量的影响(mg/g)
Table 5. Effect of soluble sugar content from Korla Fragrant Pear branches under the treatments with different concentrations of Exogenous GA3 (mg/g)
表5. 外源GA3处理对库尔勒香梨枝条可溶性糖含量的影响(mg/g)
处理枝条中可溶性糖含量变化不大,并且各处理间可溶性糖含量的差别也不大。在−24℃时,各处理的可溶性糖含量达到最高值,最高值出现在A3处理,为50.91 mg/g,比同处理的对照高15.65%,其它比对照高的有A1和B1。−26℃时,可溶性含量下降,这主要是由于低温使可溶性糖用于增加原生质的浓度,减小细胞内失水和结冰,以提高枝条组织细胞的抗寒性。各温度下各处理间的差异显著性见表5。
3.1.6. 外源GA3处理对库尔勒香梨枝条淀粉含量的影响
从表6可以看出,外源GA3处理后,随着温度的降低,各处理枝条内淀粉含量逐渐降低。在4℃至−24℃范围内,各处理内淀粉含量逐渐降低,且降低比较缓慢;在−24℃之后,除C3处理外,淀粉含量降低均在26%以上。说明在−24℃后,淀粉转化成其他小分子物质,来调节细胞内的渗透平衡,由此来提高抗寒性。其中降幅最大的是A2和A3,降幅达到45%以上。−22℃温度下,各处理间淀粉含量的差异不显著,其它处理温度下各处理之间的差异性如表6所示。
3.2. 各抗寒指标的隶属函数值及平均隶属度
为了全面利用各指标对库尔勒香梨的抗寒性进行综合评价,通过计算各综合指标的的隶属函数值来分析比较并排序(表7)。
由表7可看出,除了C1和C2处理枝条的平均隶属度小于0.4,其它处理枝条的平均隶属度均大于0.4,并且A3处理的平均隶属度最大,大大地高于其它8个处理。
根据平均隶属度的划分,从表7来看A3处理,即100 mg/L GA3一次处理枝条的平均隶属度为0.814 > 0.7,因此100 mg/L GA3一次处理为高抗,划分为抗寒Ⅰ级;50 mg/L GA3一次处理、75 mg/L GA3一次处理、50 mg/L GA3二次处理、75 mg/L GA3二次处理、100 mg/L GA3二次处理、100 mg/L GA3三次处理的平均隶属度均大于0.4,因此以上各处理均为中抗,划分为抗寒Ⅲ级;50 mg/L GA3三次处理、75 mg/L GA3三次处理的平均隶属度大于0.3,因此这两种处理为低抗,属于抗寒Ⅳ级。
4. 讨论
本试验通过对外源赤霉素(GA3)处理过的库尔勒香梨枝条进行电导率、丙二醛、脯氨酸、可溶性蛋白
Table 6. Effect of starch content from Korla Fragrant Pear branches under the treatments with different concentrations of Exogenous GA3 (mg/g)
表6. 外源GA3处理对库尔勒香梨枝条淀粉含量的影响(mg/g)
Table 7. Membership value and average membership of Korla Pear branches with different concentrations of Exogenous GA3 treatments under artificial low temperature conditions
表7. 人工低温条件下库尔勒香梨枝条外源GA3处理的隶属函数值及平均隶属度
质、可溶性糖及淀粉6大生理指标的测定,以期为库尔勒香梨的外施GA3抗寒效果的鉴定提供预测性的评价。
实验结果表明,外施不同浓度的GA3延缓了库尔勒香梨的电解质渗出率,且外施次数越多,缓解效果相对降低。随着低温胁迫的加剧,丙二醛含量总体呈现上升趋势,而−24℃至−26℃枝条中的丙二醛含量急剧上升;丙二醛是细胞膜脂过氧化作用的反映,丙二醛含量的增减是树体为适应外界环境的变化而做出的一系列的调节反应。游离脯氨酸的含量随着温度的降低平稳地增加,相对而言,外施一次GA3脯氨酸增加较大且比较平稳。本实验中可溶性蛋白质的含量随着温度的降低持续增加,含量均高于对照。各处理的可溶性糖含量均在−24℃时达到最高值,其中A3处理的糖含量最高,且差异显著(P < 0.05);淀粉含量降低最大的是A组处理;在−26℃下达到最低值,各处理间的差异显著(P < 0.05)。
植物的抗性受多种因素影响,不能单一的用某一生理指标来揭示植物抗寒的本质。各生理指标不但有各自的单方面作用,而且相互之间的作用也有重要的影响,只有对这些指标的交互作用深入综合分析,才能提高对植物抗寒性的准确鉴定。隶属函数分析提供了在多指标测定的基础上对材料进行综合评价的方法,将它应用于抗性品种的选择,更具科学性和可靠性[30] 。
本研究先用多种方法对植物生理6大指标进行抗寒性鉴定,然后用隶属函数法对其进行了抗寒性综合评价。初步研究了在人工低温胁迫下,不同浓度不同次数喷施外源GA3,库尔勒香梨树离体一年生枝条的抗寒生理指标的变化以及与抗寒性的关系。
5. 结论
综合分析各抗寒指标及平均隶属值可知,外源GA3处理叶片可以提高库尔勒香梨树的抗寒性。可初步认为,最佳GA3喷施浓度为100 mg/L,处理一次,能够抵抗−24℃的低温。但最佳喷施时间、外源激素浓度对库尔勒香梨抗寒生理的影响,以及外施激素后对内源激素的影响仍需要进一步深入研究。