1. 引言
智利竹筴鱼(Trachurus murphyi Nichols 1920)为大洋洄游鱼类,广泛分布于东南太平洋的浅水和大洋水域,常出现于沿岸和岛屿陆架、浅滩和海山的浅水区。为集群性品种,常密集成群。主要摄食桡足类、小型甲壳类、虾类、小鱼和头足类,于夏季繁殖,产浮性卵[1] 。智利竹筴鱼是重要的商业捕捞对象,其公海渔业目前置于南太平洋区域渔业管理组织(SPRFMO)管理之下。我国自2000年起开始进行智利竹莢鱼捕捞。但近年来智利竹莢鱼种群波动剧烈,产量也不稳定,因此需对智利竹莢鱼渔业现状进行评述,为进一步开展评估智利竹莢鱼资源开发潜力工作提供基础资料,也对指导渔业企业开展生产安排提供参考信息。
2. 渔场情况
智利竹筴鱼分布区域广泛,其分布区东起智利、秘鲁和厄瓜多尔专属经济区,向西延伸至新西兰外海,覆盖了78˚W~177˚W间狭长的南太平洋水域。智利竹筴鱼的分布区域,也被称为“竹筴鱼带”(图1) [2] 。
智利竹筴鱼呈现明显的季节性洄游模式。春季,竹筴鱼离岸向大洋洄游,主要分布在32˚S~40˚S之间、90˚W以西的智利外海水域;夏末,因沿岸海域饵料丰富,向岸索饵洄游;秋、冬季,则成群聚集在智利专属经济区内[3] 。
智利竹筴鱼的空间分布,因其发育程度不同而有所差异。成熟的竹筴鱼产卵群体,主要集中于35˚S~40˚S范围内,离岸向西达90˚W的产卵场。而成鱼的索饵场主要分布在智利沿海海表温低于15℃中–南部水域(33˚S~40˚S),幼鱼发育成熟后会补充入该区域。育幼场分布在15℃等温线以北温暖的大洋性水域和沿岸水域。总体来说,智利沿岸对智利竹筴鱼的育幼和补充至关重要[2] [4] 。
化成君等[5] 根据2005~2008年3~11月份我国在东南太平洋的智利竹筴鱼大型拖网渔捞日志资料,获得了智利竹筴鱼公海渔场的年际和季节变化规律(图2)。
从产量重心的季节变化来看,智利竹筴鱼的公海渔场分布具有明显的季节变化,秋季渔场分布纬度处于最南端,变化范围较小,主要分布于80˚W~87˚W、41˚S~45˚S范围内,冬季渔场纬度逐渐北移,春季渔场处于最北端,变化范围最大,反映春季处于渔汛末期,鱼群分散索饵,意味着渔场快要结束[5] 。
智利竹莢鱼渔场的形成与西风漂流与秘鲁寒流关系密切。西风漂流与秘鲁寒流向温暖海域输送的冷水,形成智利、秘鲁外海强盛的涌升流,将大量硝酸盐、磷酸盐等营养物质带到海洋中上层。这促进了浮游生物大量繁殖,为竹莢鱼提供了丰富的饵料。因此,西风漂流带成为竹筴鱼分布的南部屏障(45˚S左右),秘鲁寒流控制着智利竹筴鱼在东西方向上的分布[1] 。
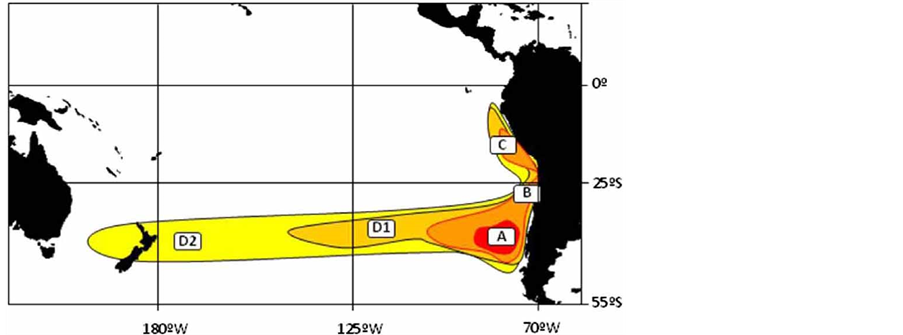
Figure 1. Distribution of Chilean jack mackerel, A, south-central Chilean stock; B, north Chilean stock; C, Peru stock; D1, south-central Pacific stock; D2, south-west Pacific stock [2]
图1. 智利竹筴鱼分布概略图,A为智利中南部智利竹筴鱼群;B为北部智利竹筴鱼群;C为秘鲁群;D1为中南部太平洋群;D2为西南部太平洋群[2]
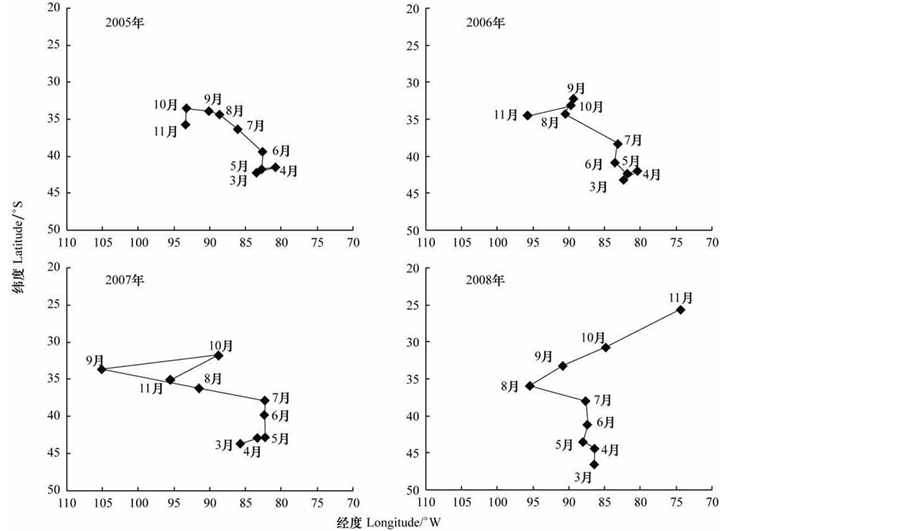
Figure 2. Yield center of Chilean jack mackerel fishing ground in Mar. to Nov. from 2005 to 2008 [5]
图2. 2005~2008年3~11月份智利竹筴鱼渔场产量重心的时空变化[5]
综合现有资料,智利竹莢鱼渔场的分布,受到其洄游路线、水温、季节与洋流体系的影响。安排生产时应注意洋流与温度变化,春季至秋季作业区域应由北向南,冬季则向北作业。此外,应注意保护沿岸温暖水域的育幼场。
3. 产量与资源波动
SPRFMO (2014) [6] 调查报告显示,上世纪70年代以后,智利竹筴鱼的资源量经历了大幅波动(图3)。在80年代晚期,智利竹筴鱼的补充和产卵群体生物量曾达到历史最高峰,但自90年代之后经历了迅速下滑。本世纪初,智利竹筴鱼资源量曾有恢复迹象,但随后继续衰退。
智利竹筴鱼捕捞量与其种群资源量变化趋势较为相似。自1950年起,智利开始在其专属经济区内开发竹筴鱼资源,其渔获量占据了全球竹筴鱼渔获量的绝大部分。而东南太平洋公海的智利竹筴鱼渔业,由前苏联船队于1978年开始开发。1991年以前,智利竹筴鱼产量一直稳步上升,其中前苏联捕获了1.3 × 107 t智利竹筴鱼。但此后,随着前苏联的解体及其他原因,智利竹筴鱼渔获量经历了大幅度波动,在1995年达到4.95 × 106 t高峰后便迅速下降,到1999年进入低谷,年产量仅为1.42 × 106 t,之后又有所回升,到2001年恢复到2.52 × 106 t水平,但自2008年以来,产量逐渐下降,到2011年,产量仅为0.63 × 106 t (图4) [6] 。
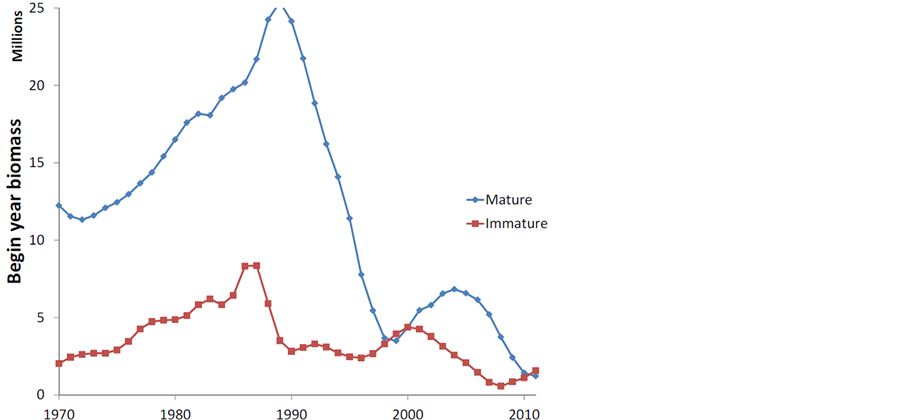
Figure 3. The biomass of Chilean jack mackerel from 1970 to 2011[6]
图3. 1970~2011年智利竹莢鱼生物量波动趋势[6]
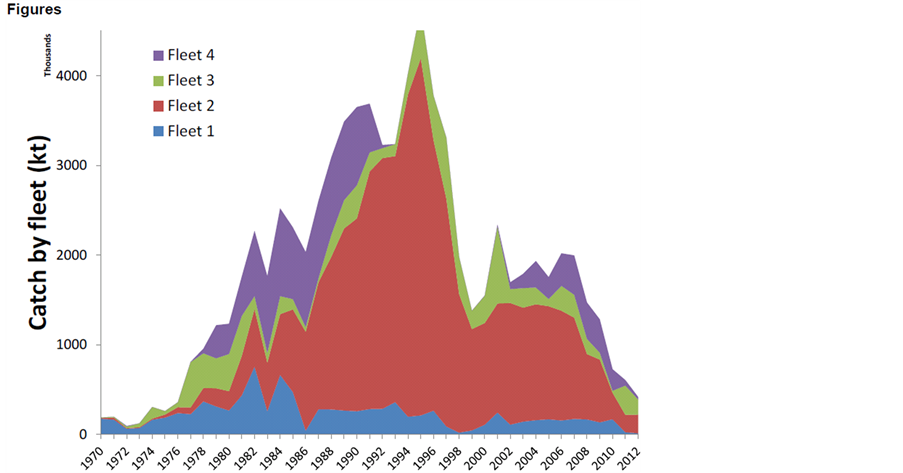
Figure 4. Annual catch of Chilean jack mackerel by fleets (Fleet 1: North of Chilean coast, Fleet 2: South-center of Chilean coast, Fleet 3: Coast and off shore of Peru, Fleet 4: International fishery) [6]
图4. 智利竹荚鱼各组船队历年产量(其中船队分别为:1,智利北部;2智利中部和南部;3秘鲁沿岸及外海;4公海渔业) [6]
中国渔船在在2001~2002年时,开始进行智利竹荚鱼探捕,此阶段以寻找渔场为主,受捕捞能力限制,产量和CPUE都不高。随张对渔场情况的逐渐熟悉及捕捞能力的提升,中国船队的智利竹筴鱼渔获量在2004~2007年间处于高位。至2009年,中国船队的CUPE达到最高值,但产量却持续下滑,至2012年产量下降至最低不足2万吨,但CPUE止住下滑趋势(图5)[7] 。
4. 波动产生的原因
造成这种总产量大幅度起落的原因,可能是多方面的。渔业生产方面,前苏联加盟国大批船只退出了对智利竹筴鱼的捕捞,造成了90年代初产量的大幅度下降。但是近年来,虽然捕捞努力量不断增加,东南太平洋智利竹筴鱼的产量还是呈下降趋势。
近10余年间,智利外海竹筴鱼的种群特征发生了变化。从渔获量最高的智利中部与南部海域各年龄段渔获比例来看,21世纪以前,主要渔获目标为较低龄个体,但自本世纪以来,高龄个体逐渐成为主要渔获物,低龄个体极少,在2007~2008年间达到高峰,从侧面反映出,补充群体在这些年份的衰退。此后低龄个体所占比例略有回升(表1)[6] 。
上世纪90年代中后期,低龄个体渔获量较高,一方面表示与补充群体生物量仍比较充分有关,另外也与对智利竹筴鱼开发仅剩智利等沿海国家有关(图4)。由于智利竹筴鱼最大的育幼场即在此区域,因此有大量低龄鱼在该水域被捕获。
而90年代,智利在本国专属经济区内的捕捞量达到高峰,随后该国捕捞产量也开始迅速下降。这从侧面反映出,低龄种群经受过高的捕捞压力,可能对育幼场种群造成了破坏。而研究指出,智利竹筴鱼生命周期大约为15年,3龄鱼、体长30 cm左右性成熟[8] 。育幼场的破坏,导致部分竹莢鱼在未能达到性成熟即被捕获,影响了种群的补充。这可能是随后公海渔业重新开始开发该资源后始终无法达到苏联时代的最高产量的原因之一。
从智利竹筴鱼鱼卵与仔幼鱼分布来看(图6) [3] ,在2003年之后,鱼卵与仔幼鱼密度大幅下降,虽然在2004年有小幅的上升但随后仍然呈现剧烈的下降趋势。而2007年之后智利竹筴鱼产量、资源量大幅度下降,也与2003年之后鱼卵与仔幼鱼发生量下降,从而导致补充群体受限,有重要关系。但值得注意的是,2004~2006年是21世纪以后智利竹筴鱼资源量和产量的高峰(图3),但鱼卵与仔幼鱼发生量却陷入低谷。
这一不依赖亲体密度的悖反现象暗示,对于智利竹筴鱼资源动态的分析,除育幼场破坏之外,还需
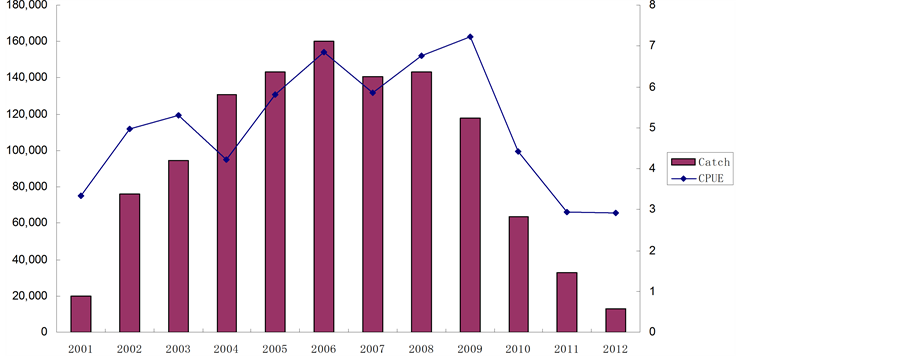
Figure 5. Chilean jack mackerel catch and CPUE of Chinese fleet
图5. 中国渔船2001~2012年智利竹筴鱼产量与CPUE

Table 1. Age compose of catch from south-center of Chilean coast (%)
表1. 智利中部与南部海域各年龄段渔获所占比例(%)
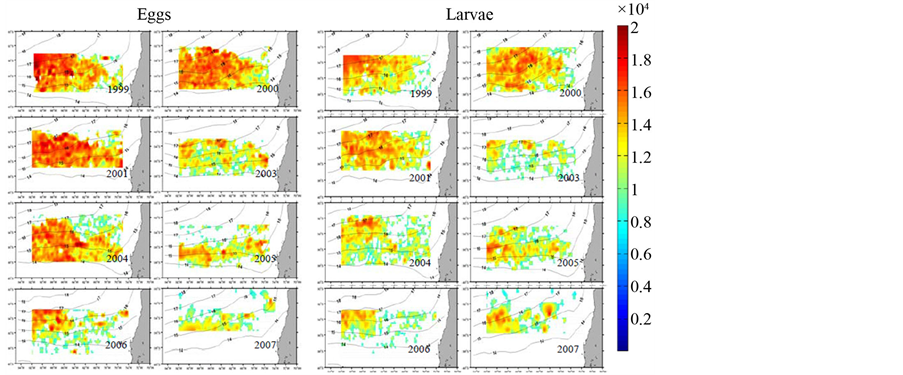
Figure 6. Distribution of Chilean jack mackerel eggs and larva from 1999 to 2007 [3]
图6. 智利竹筴鱼1999~2007年鱼卵与仔幼鱼分布图[3]
要充分考虑气候变迁导致的环境改变因素对资源量的影响。例如,1997~1998年出现强埃尔尼诺现象对秘鲁外海竹筴鱼分布变化的影响已有研究发表。气候的改变通过环境条件和饵料的变化影响了智利竹筴鱼的分布[4] 。
因此,从现有研究资料来看,造成智利竹莢鱼资源与产量大幅波动的主要原因包括:(1) 捕捞努力量投入的大幅波动,(2) 育幼场曾经遭受破坏,(3) 气候变迁造成的影响。这提示我们,在评估智利竹莢鱼资源开发潜力时,除依据其渔业捕捞生产动态之外,还需要考虑到其世代补充,也要注意气候变化对渔场环境的影响。

Figure 7. Dynamic of Chilean jack mackerel fishing mortality vs. biomass [6]
图7. 智利竹筴鱼捕捞历年死亡率与生物量变化[6]
5. 资源量现状与开发潜力
SPRFMO科学委员会报告[6] 对智利竹筴鱼资源状况进行了综合评价。报告认为,自1991年之后,随着苏联对智利竹筴鱼渔业开发力度萎缩,捕捞死亡率有短暂下降,但捕捞努力量早已维持在最大可持续捕捞死亡率之上,因此自93年之后,随着智利的捕捞强度急剧增加,智利竹筴鱼的生物量也迅速下降,1995年以后,开始低于最大可持续生物量。98年后捕捞死亡率出现大幅下降后,生物量仍未能恢复(图7)。
21世纪以来,智利竹筴鱼生物量始终低于最大可持续生物量,而在2012年之前,捕捞死亡率则一直高于最大可持续捕捞死亡率。随着2010年之后,许多渔船退出智利竹筴鱼渔业,而智利和秘鲁先后加大了针对智利竹筴鱼的捕捞死亡率限制,智利竹筴鱼资源已经停止了下降趋势,近年来捕捞努力量大幅度削减,导致智利竹筴鱼捕捞死亡率目前低于最大可持续捕捞死亡率,暂时脱离了捕捞过度状态。但由于智利竹筴鱼种群2007年以来的持续快速衰退,虽经过近2年来的小幅度恢复仍然低于最大可持续生物量,因此当前本资源处于过度开发状态(图7) [6] 。
综合上述材料,智利竹莢鱼资源量目前仍然未能恢复到足以维持最大可持续产量的水平上。但是,经历了近年来持续的严格管理及捕捞努力量下降之后,其捕捞死亡率已经降至较低水平,承受的捕捞压力也较轻,因此具备了种群恢复的可能。而中国渔船在公海的生产数据显示,智利竹莢鱼的CPUE在2012年已经止住了下滑趋势,也从侧面反映出该资源有恢复的迹象。而近年来3龄以下智利竹莢鱼所占渔获比例已经控制在一个很低的水平,使育幼场得到了保护,补充群体自2009年起持续增长。因此从世代补充关系角度分析,智利竹莢鱼资源在未来数年内具备恢复的条件。此外,气候的波动是预测智利竹莢鱼种群动态的重要不确定性因素,所以深入研究智利竹莢鱼对气候波动的响应十分必要。
基金项目
本综述得到国家科技支撑计划项目(2013BAD13B05-3)与2013年远洋渔业资源调查和探捕项目的“东南太平洋公海竹筴鱼资源”专题研究资助。

NOTES
*通讯作者。