1. 引言
工业企业的临海布局、人类活动的增加,远超海洋环境容量,造成海洋滩涂重金属含量超标,破坏海洋湿地生态系统,影响海产品品质,严重制约当地的渔业经济,也不利于滨海经济的可持续发展 [1] [2] 。Pb2+、Cd2+是动植物生长的非必需元素 [3] ,镉会导致细胞死亡,植物黄化枯萎 [4] 。在铅胁迫时,即使很低的浓度,植物组织也会受到损害,如微核,DNA破坏 [5] ,膜通透性的改变 [6] 和部分酶活性降低 [7] 等。此外,铅对镉在植物体内的积累有增强效果 [8] 。植物对铅和镉累积,也会刺激超氧自由基产生,造成植物氧化应激 [9] ,使膜脂过氧化,引起超氧化物歧化酶(Superoxide Dismutase, SOD)活性、丙二醛(Malondialdehyde, MDA)含量、脯氨酸(Proline, Pro)含量等生理生化指标发生变化 [10] [11] [12] 。
碱蓬(Suaeda salsa)属藜科,是本土一年草本真盐生植物,常见于盐碱地及沿海滩涂,在新疆、蒙、甘、陕、鲁、浙、晋、辽等省区均有分布。碱蓬3~6月出苗,7~8月为花期,9~10月为结实期,11月为种子枯熟期 [13] 。由于碱蓬耐盐性强,又为土著植物,从而受到盐碱湿地修复领域的广泛关注,对其重金属累积能力 [14] 、有机物吸收能力 [15] 以及对氮磷的吸收与再分配作用 [16] 等都有相关的研究报道,在一些沿海滩涂地区盐生植物碱蓬作为优势植物,可明显吸收盐渍土壤中的Cd、Pb等重金属 [17] ,被誉为盐碱地修复的先锋植物 [18] 。本文研究并比较了不同生长时期的碱蓬(幼苗期和花期),在Pb2+、Cd2+单一及复合胁迫下的生理生化响应,旨在为碱蓬的生长代谢过程中长期适应和抵御Pb2+、Cd2+胁迫的损伤机制,及其修复重金属污染的高盐碱滩涂应用等提供理论依据。
2. 材料与方法
2.1. 试剂与材料
盐地碱蓬(采集于杭州湾庵东湿地);塑料花盆(直径16 cm,高16 cm);蛭石(φ = 2~4 mm,填充花盆至2/3体积)、海水(取自象山港)。蛭石和海水中的Pb2+、Cd2+背景值如表1。
2.2. 试验方法
根据《土壤环境质量标准》 (GB15618-2008)、土壤背景值及工业区附近土壤中的Pb2+、Cd2+浓度,进行完全交互实验设计,共16个处理组,每个处理4盆重复,如表2所示。
实验分别于当年5月(幼苗期),7月(花期)开始,时间均为60天,Pb2+和Cd2+以Pb(NO3)2∙6H2O、Cd(NO3)2∙4H2O配成溶液形式加入,于种植前2周平衡固定、装盆,其中,不添加任何重金属的为对照组。当年5月,采集株高相近的幼苗,洗净根部泥沙,种于上述盆中,每盆2棵,将蛭石填入花盆中,占盆体积2/3,以Hoagland营养液:海水 = 1: 1 (盐度约为1.6%)定期浇灌,室外自然光照下培养,以Hoagland营养液:海水 = 1: 1 (盐度约为1.6%)定期浇灌,既能保持基质湿润,还能满足碱蓬对盐分的需求,并逐步适应滨海环境中的土壤盐度。参照文献 [19] ,在幼苗期、花期分别检测株高,以盆为单位,采集每株主干上相同高度、成熟度一致且生长状况相近的绿色叶片,测量其超氧化物歧化酶(SOD)活性(氮蓝四唑法) [20] 、丙二醛(MDA)含量(硫代巴比妥酸法) [21] 、脯氨酸(Pro)含量(酸性茚三酮比色法) [22] ,每组重复测量3次。数据采用SPSS 20组间方差分析、Graph Pad Prism 5作图,结果以均值±标准差表示。
3. 结果与分析
3.1. Pb2+、Cd2+胁迫对碱蓬株高的影响
由图1可知,幼苗期,随单一Cd2+浓度增加,促进碱蓬生长,使株高显著高于对照组(P < 0.05),浓度继续增大,抑制碱蓬生长,使株高显著小于对照组(P < 0.05),且浓度越大,抑制作用越强。单一Pb2+浓度增加对株高起促进-抑制-促进作用,加入5 mg∙kg−1的Cd2+使其逆转为抑制-促进-抑制作用,分别以20, 50 mg∙kg−1的Cd2+与Pb2+ (除200 mg∙kg−1外)复合,均抑制碱蓬生长。生长至花期,对照组株高最大

Table 1. Background values of Pb2+, Cd2+ in the vermiculite and sea water
表1. 蛭石和海水中的Pb2+、Cd2+背景值
Table 2. Experimental factor-level table
表2. 实验因素-水平表

Figure 1. Effects of Pb2+, Cd2+ single and combined stress on the height of Suaeda salsa. Note: Cd2+ 0 mg∙kg−1; Cd2+ 5 mg∙kg−1; Cd2+ 20 mg∙kg−1; Cd2+ 50 mg∙kg−1
图1. Pb2+、Cd2+单一及复合胁迫对碱蓬株高的影响。注:Cd2+ 0 mg∙kg−1;Cd2+ 5 mg∙kg−1;Cd2+ 20 mg∙kg−1;Cd2+ 50 mg∙kg−1
(P < 0.05),单一Pb2+、Cd2+胁迫使株高随浓度增加而减小,但500 mg∙kg−1 Pb2+对株高的抑制作用小于200 mg∙kg−1的Pb2+胁迫。幼苗期与花期,不同浓度Cd2+与200 mg∙kg−1的Pb2+复合胁迫,株高均大于相应的单一Pb2+胁迫,表现为拮抗作用,使毒性减小;与500, 20 mg∙kg−1的Pb2+复合胁迫,株高均小于相应的单一Pb2+胁迫,表现为协同作用,其中,500 mg∙kg−1的Pb2+与50 mg·kg−1的Cd2+复合胁迫下株高最小,协同作用显著(P < 0.05),毒性加强,抑制生长,这表明碱蓬的两个生长时期下,Pb2+、Cd2+的相互作用属性不变。重金属影响植物生长,低浓度促进植物生长,高浓度抑制生长,甚至引起植物死亡 [23] [24] 。其中,Pb2+对植物的毒害表现为矮化植物,影响矿质元素的吸收;Cd2+则能与蛋白质相互作用,对细胞产生毒性 [2] 。试验结果中花期对照组株高最高,表明随Pb2+、Cd2+胁迫时间的延长,重金属对碱蓬主要表现为毒害作用,抑制生长。
3.2. Pb2+、Cd2+胁迫对碱蓬SOD活性的影响
SOD活性是植物抗氧化保护系统的第一道防线,主要是清除活性氧,使细胞膜免受氧自由基的伤害,当受到一定范围内的胁迫时,活性会有所提高,增加植物的抗逆能力,其水平高低是衰老与死亡的直观指标 [25] 。由图2可知,幼苗期,单一Cd2+ (5 mg∙kg−1, 50 mg·kg−1)胁迫使SOD活性显著高于对照组,说明一定浓度的Cd2+能诱导SOD合成保护机体;单一Pb2+胁迫以及与低浓度的Cd2+ (5 mg·kg−1, 20 mg·kg−1)复合胁迫使SOD活性随Pb2+浓度增加先升高后降低,峰值分别出现在20 mg·kg−1、200 mg·kg−1的Pb2+处;高浓度的Cd2+ (50 mg·kg−1)胁迫下,SOD活性随Pb2+浓度的增加一直下降。Pb2+与Cd2+复合胁迫,SOD活性均显著小于对应的单一Pb2+胁迫(P < 0.05),说明在复合胁迫中,Cd2+对碱蓬SOD活性影响较大,会阻碍SOD的合成。
花期碱蓬SOD活性约300 U·g−1,极显著低于幼苗期(P < 0.01),表明碱蓬已开始进入衰老期,符合生长周期规律。对照组SOD活性最大(P < 0.05),Pb2+、Cd2+单一及复合胁迫的SOD活性均显著小于对照组(P < 0.05);且SOD活性变化较小;SOD活性在500 mg∙kg−1 Pb2+ + 20 mg·kg−1 Cd2+胁迫下最小。这可能是由于胁迫时间较长,植物吸收累积了一定重金属,阻碍SOD合成;也可能是与植物进入衰老期相关,说明碱蓬在花期自我调节SOD活性来适应外界胁迫的能力有限。
3.3. Pb2+、Cd2+胁迫对碱蓬Pro含量的影响
Pro是植物体内普遍存在的一种逆境保护性物质,有助于抵御逆境胁迫,植物能通过提高其含量调节渗透平衡,保护细胞结构和功能的完整性,其含量在一定程度上反映了植物的抗逆能力 [10] [26] 。由图3
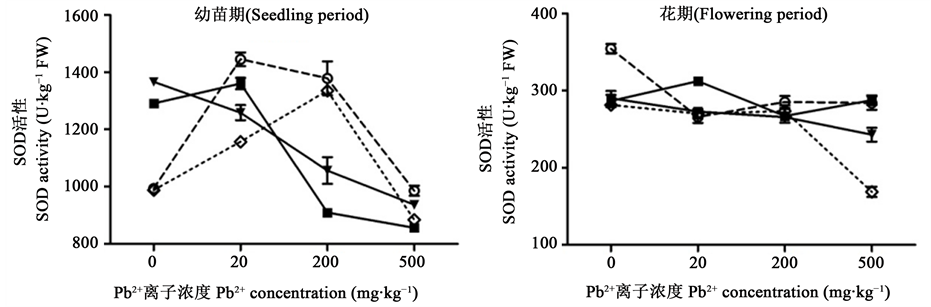
Figure 2. Effects of Pb2+, Cd2+ single and combined stress on the SOD activity of Suaeda salsa. Note: Cd2+ 0 mg∙kg−1; Cd2+ 5 mg∙kg−1; Cd2+ 20 mg∙kg−1; Cd2+ 50 mg∙kg−1
图2. Pb2+、Cd2+单一及复合胁迫对碱蓬SOD活性的影响。注:Cd2+ 0 mg∙kg−1;Cd2+ 5 mg∙kg−1;Cd2+ 20 mg∙kg−1;Cd2+ 50 mg∙kg−1

Figure 3. Effects of Pb2+, Cd2+ single and combined stress on the Pro content of Suaeda salsa. Note: Cd2+ 0 mg∙kg−1; Cd2+ 5 mg∙kg−1; Cd2+ 20 mg∙kg−1; Cd2+ 50 mg∙kg−1
图3. Pb2+、Cd2+单一及复合胁迫对碱蓬Pro含量的影响。注:Cd2+ 0 mg∙kg−1;Cd2+ 5 mg∙kg−1;Cd2+ 20 mg∙kg−1;Cd2+ 50 mg∙kg−1
可知,幼苗期,单一Pb2+、Cd2+胁迫使Pro含量随重金属浓度增加先升高后降低;复合胁迫时,Pro含量随Pb2+浓度增加而下降;Pro含量均比对应的单一Pb2+胁迫低(P < 0.05),说明两种重金属复合作用对碱蓬Pro含量影响较大,为协同毒害作用。除200 mg∙kg−1 Pb2+ + 50 mg∙kg−1 Cd2+、500 mg∙kg−1 Pb2+ + 5 mg∙kg−1 Cd2+外,其余浓度复合胁迫时,花期碱蓬Pro含量显著高于对照组(P < 0.05),说明碱蓬能通过提高渗透平衡保护细胞结构和功能完整,有助于抵御逆境胁迫,但在高浓度Pb2+、Cd2+复合胁迫下,Pro的自我调节能力会受到限制。中高浓度Cd2+ (20, 50 mg∙kg−1)胁迫使Pro含量随Pb2+浓度增加而下降;单一Pb2+及Pb2+与低Cd2+ (5 mg∙kg−1)复合胁迫,则使Pro含量随Pb2+浓度增加先升高后下降,峰值均出现在200 mg∙kg−1的Pb2+处,表明碱蓬对外界胁迫响应强烈,这可能与200 mg∙kg−1的单一Pb2+胁迫抑制碱蓬生长相关。
3.4. Pb2+、Cd2+胁迫对碱蓬MDA含量的影响
MDA是植物细胞膜脂过氧化产物之一,能与细胞内各种成分发生反应,引起各种酶和膜的严重损伤,其含量高低可以反应膜脂过氧化作用强弱及质膜损坏程度 [27] 。由图4可知,幼苗期,加入低浓度的Cd2+ (≤20 mg∙kg−1)使碱蓬MDA含量随Pb2+浓度增加而下降,加入高浓度的Cd2+ (50 mg∙kg−1)使碱蓬MDA
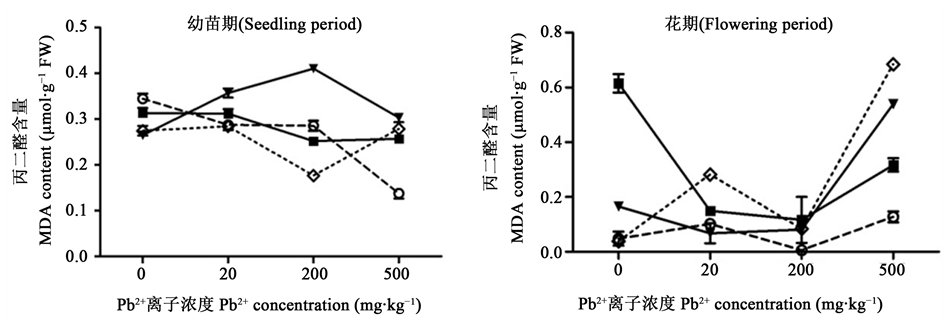
Figure 4. Effects of Pb2+, Cd2+ single and combined stress on the MDA content of Suaeda salsa
图4. Pb2+、Cd2+单一及复合胁迫对碱蓬MDA含量的影响
含量随Pb2+浓度增加先升高后下降。其中,高浓度的Pb2+ (500 mg∙kg−1)与任意浓度的Cd2+复合,MDA含量显著大于单一的Pb2+胁迫(P < 0.05),表明高Pb2+胁迫下,加入不同浓度的Cd2+均能加剧碱蓬质膜损伤;高浓度Cd2+(50 mg∙kg−1)与Pb2+复合的MDA含量高于低Cd2+ (5 mg∙kg−1)与Pb2+复合,说明幼苗期在高浓度的Cd2+胁迫下时,质膜损伤最为严重,在200 mg∙kg−1 Pb2+ + 50 mg∙kg−1 Cd2+时,MDA含量达到最大。花期,碱蓬MDA含量基本高于对照组,500 mg∙kg−1 Pb2+与20 mg∙kg−1 Cd2+复合胁迫下的MDA含量最大;200 mg∙kg−1、500 mg∙kg−1的Pb2+与任意浓度的Cd2+复合,均显著大于对应的单一Pb2+胁迫(P < 0.05),表明在单一和复合胁迫下,质膜都受损严重,加入Cd2+使其损伤更大,表现为协同毒害作用。
4. 讨论
大部分研究表明 [23] [25] [27] (周玉卿等,2012;徐学华等,2009;郭天荣等,2015),低浓度的重金属对植物有促进作用,高浓度则为抑制作用,甚至导致死亡。本试验中,幼苗期碱蓬在单一Cd2+胁迫下符合这一规律;在单一Pb2+胁迫下,低浓度(20 mg·kg−1)的Pb2+促进生长,较高浓度(200 mg·kg−1)的Pb2+抑制生长,更高浓度(500 mg·kg−1)的Pb2+反而促进碱蓬生长,出现异常,这可能是由于碱蓬对高Pb2+存在一种规避机制,与何洁 [28] 研究翅碱蓬对单一铅的累积吸收结果类似;但随碱蓬继续生长,胁迫时间延长,花期碱蓬在各个浓度胁迫下的株高均显著小于对照组(P < 0.05),抑制作用显著,这可能是由于碱蓬体内累积了一定量的重金属,阻碍碱蓬生长。单一胁迫与复合胁迫相较,任意浓度的Cd2+与200 mg·kg−1的Pb2+复合胁迫,株高均显著大于相应的单一Pb2+胁迫(P < 0.05),为拮抗作用;与500、20 mg·kg−1的Pb2+复合胁迫,株高均显著小于相应的单一Pb2+胁迫(P < 0.05),为协同作用,并非两种重金属的复合毒害作用一定强于单一重金属的毒害,这与刘大林 [10] 以Pb2+、Cd2+胁迫饲用高粱、郭天荣 [27] 以Pb2+、Cd2+胁迫空心菜时,复合胁迫较单一胁迫会加剧对植物的毒害作用的结果不同,可能原因是与植物本身的调节方式有关,或许又与重金属在植物体内存在的部位、形式、累积量有一定联系,有待于进一步研究。
逆境胁迫对植物的损伤常常是由于产生过多的体内活性氧(ROS),从而导致膜脂受损,细胞功能丧失。植物往往能有限地在体内合成一些物质来进行自我调节,以适应外界环境 [25] 。SOD活性在重金属胁迫下,往往有3种表现形式 [24] :一是随浓度的增加而增加;二是随浓度增加先升高后下降;三是随浓度的增加而下降,本试验的SOD活性符合后两种情况。单一Pb2+胁迫以及与20 mg∙kg−1的Cd2+复合胁迫,SOD活性均随Pb2+增加先升高后降低,而与5, 50 mg∙kg−1的Cd2+复合胁迫,SOD活性随Pb2+浓度增加而降低,这可能是由于Cd2+浓度的增加使SOD调节机制受损,但其具体受损机理需进一步研究。
幼苗期,碱蓬在单一Pb2+、Cd2+胁迫下,Pro含量随重金属浓度增加先升高后降低;与Cd2+复合胁迫时,Pro含量随Pb2+浓度增加而降低;花期碱蓬的Pro含量基本高于对照组,说明幼苗期碱蓬的Pro调节能力较弱,而在花期Pro调节作用变得较为明显。在单一重金属胁迫时,Pro与SOD相比较,Pro在遇到重金属胁迫时,其响应较慢,在重金属浓度升高后才逐渐产生且产生量较SOD少,因此其调节存在先后,SOD为先,Pro为后。
幼苗期的MDA含量高于花期,随Pb2+浓度增加呈无序变化,与孙永林 [12] 在研究Pb2+对黄瓜的生理影响中,随Pb2+浓度增加MDA含量也增加的结果不同,这可能是碱蓬对Pb2+存在独特的调节机制。
综上所述,幼苗期,SOD活性能迅速对重金属胁迫响应,Pro在花期其渗透调节作用变得较为明显能抵御外界重金属胁迫,MDA含量呈无序变化,代表机体膜质受损状况,其无序变化随外界重金属胁迫及碱蓬体内Pro、SOD活性等组成的调节系统的动态变化而变化。因此,碱蓬在抵御外界重金属胁迫的整个过程中,会启动SOD、Pro两种物质协同抵抗,各自的调节能力有限,且存在先后之分,并能通过MDA含量实时表征植物的受损情况。
5. 结论
(1) Pb2+、Cd2+单一及复合胁迫均对碱蓬有毒害作用,且时间延长毒害作用增加,表现为抑制碱蓬生长;(2) SOD和Pro对重金属调控起重要作用,SOD响应为先,Pro为后;(3) 无论在幼苗期还是花期,当200 mg∙kg−1 Pb2+与20、50 mg∙kg−1 Cd2+复合胁迫时,重金属之间表现为拮抗作用,其他Pb2+浓度与Cd2+复合时均为协同作用。总之,碱蓬对重金属胁迫有独特的调节机制和较强的耐受性,且应对重金属复合胁迫的耐受性更强,因此碱蓬对于滨海滩涂铅、镉重金属污染修复有极大的应用潜力。
基金项目
浙江省自然科学基金项目[The Project Supported by Zhejiang Provincial Natural Science Foundation of China] (LY16C150002);浙江省公益性技术应用研究计划项目[The Research Project on nonprofit technology of Zhejiang Province] (2015C32035);浙江省重中之重学科自设课题[Zhejiang Provincial Top Key Discipling Under Grant] (ZS2016011);浙江万里学院引进人才重点项目[Key project of Zhejiang Wanli University talents] (1741000530);国家级大学生创新创业训练计划项目[National Training Program of Innovation and Entrepreneurship for Undergraduates](201610876028)资助。
NOTES
*通讯作者。