1. 引言
植物通过光合作用将太阳能、二氧化碳和水合成有机物质,构成世界所有生物的物质基础。所以植物的分布决定以植物为食的生物的分布,甚至也可能会影响取食植食性生物的肉食性生物的分布。因此研究植物的分布格局将会对其他生物分布格局的理解具有重要意义。
双子叶植物是植物王国中进化程度最高、多样性和生物量最大的一类生物。种类占植物总种类的3/4,它的分布特征对整个植物界具有重要代表性。
法国博物学家布丰(G. Buffon)于18世纪中期拉开生物地理学的序幕 [1] ,进入19世纪,德国人洪堡(A. von Humboldt)奠基了植物地理学科,他由于1799~1804在南美洲的考察而成名。他观察到山上的植物依照高度不同而分成不同的植物带。他相信世界分成许多个自然地区,各有它自己的独特的动植物集群。他于1805年开始出版30卷的系列专著,详细介绍他在南美洲的植物学观察 [2] 。
瑞士植物学家德康多勒(A. de Candolle)迅速发展了洪堡的工作,他认为植物由于风、水、或动物的携带而扩散,直到海洋、高山、沙漠的阻隔而停止,也可能由于与其竞争的其它植物的存在而停止扩散。不同的自然区域有自己的特有植物类群。他拟就了一个新词“endemic”(特有的),并界定了20个这样的地区,包括18个大陆地区,2个岛屿组群 [3] 。
半个多世纪后,也是德国的植物学家恩格勒(A. Engler),用地图详细界定了4个植物“界”的边界,“泛北极界”包括整个北温带、北寒带;“古热带界”从非洲一直延伸到太平洋岛屿;“南美洲界”包括中美地区及南美洲;“古大洋洲界”包括澳大利亚大部、新西兰南部、非洲南端及南美洲南端 [4] 。20世纪又经多次的修改 [5] [6] [7] [8] [9] ,将古大洋洲界分作澳洲界、好望角界、南极界。2001年英国学者Cox提出新的意见,撤掉好望角界及南极界,有关地区归入自己所属大陆;将古热带界分成非洲界及印度–太平洋界 [10] 。至此,全球植物区系分作5个界:泛北极界、非洲界、印度–太平洋界、澳洲界、新热带界。2011中国学者吴征镒院士提出新设古地中海植物界及东亚植物界 [11] 。
由定性方法得出的这些区划方案固然有其合理的科学内核,但也无须讳言,在分布区划分的标准及分界线的确定上难免会有失衡之处。生物地理学与其它自然学科一样,都需要由数学武装,才能真正成熟 [12] 。100多年来研究生物地理区划的相似性公式虽然多达40余个 [13] ,但似乎没有得到广大生物学者的接受和赞誉 [11] [14] ,至今还无人对全球植物分布进行定量分析的尝试,以致使地理区划成为阻碍生物地理学发展的幽灵 [15] 。我们从改进分析方法入手,突破二元比较的禁锢,提出一个新的相似性通用公式(similarity general formula, SGF) [16] 和一个新的多元相似性聚类分析法(multivariate similarity clustering analysis method, MSCA) [17] 。并在不同地域范围、不同生物类群得到验证 [18] - [25] 。本研究将对世界双子叶植物进行分析,试图为支持或修订世界植物地理区划方案提供定量依据。
2. 材料和方法
2.1. 生物类群
本研究不包括化石种类,共涉及49目366科20,809属,涵盖416,483种(表1)。分布资料主要来源于分类学家的专著 [26] - [43] 、专业性网站汇总的数据库 [44] - [49] ,也会补充一些不断发表的新资料。为了提高资料的利用率及保持聚类图的清晰度,我们以“属”作为基础生物单元(basic biologicalunit, BBU)。
2.2. 基础地理单元划分
我们不主张按照经纬度将世界陆地划分为成千上万个栅格。因为生物分布资料是生物学家长期积累的结果,不是按栅格调查的,也不可能按栅格去调查。栅格与栅格之间在调查的人力、物力、时间不可能完全一致,这样会不可避免地造成人为的差异,从而影响我们要探究的自然的差异。
按照地形、气候等生态条件和生物分布资料的详略程度,本研究把全球陆地(除南极洲)划分为67个基础地理单元(basic geographical unit, BGU) (图1)。作为聚类分析与地理区划的基础。其中以平原为主的BGU有21个,以丘陵为主的BGU有11个,以山地为主的BGU有12个,以高原为主的BGU有11个,以荒漠为主的BGU有5个,岛屿型的BGU有7个。有27个BGU处在热带,有34个BGU地处温带,有6个BGU的地域跨入寒带。
2.3. 构建数据库
用微软Access构建数据库。将各个BGU作为各列,将各个BBU作为各行。将一个属分布的行政区域记录转化为BGU记录录入数据库中,有分布记“1”,无分布不记,这些基础分布记录(Basic Distributional

Table 1. The class Magnoliopsida for analysis
表1. 供分析的世界双子叶植物
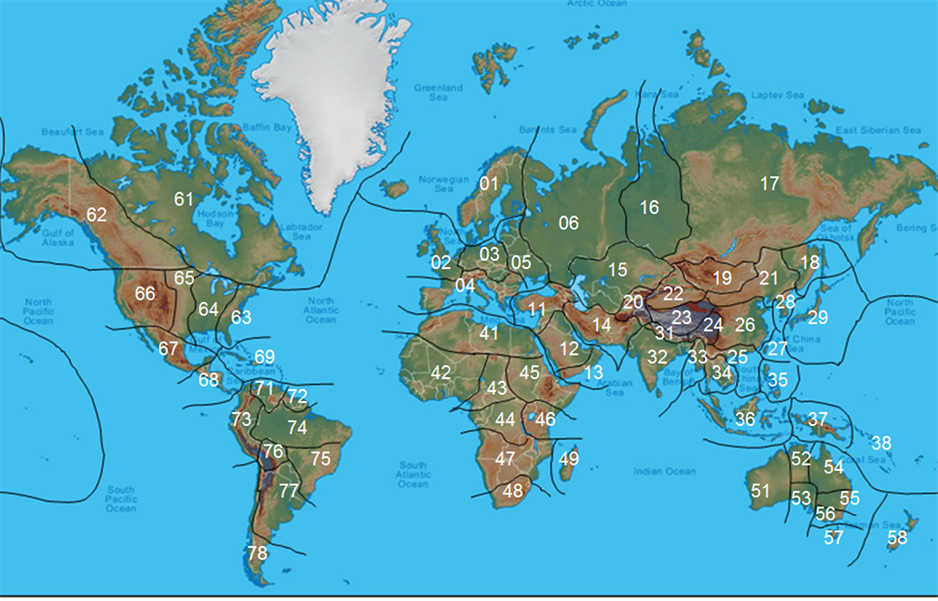 01 北欧Northern Europe, 02 西欧Western Europe, 03 中欧Central Europe, 04 南欧Southern Europe, 05 东欧Eastern Europe, 06 俄罗斯欧洲部分European Russia, 11 中东Middle East, 12 沙特阿拉伯Saudi Arabia, 13 也门与阿曼Yemen and Oman, 14 伊朗高原Plateau of Iran, 15 中亚Central Asia, 16 西西伯利亚Western Siberia, 17 东西伯利亚Eastern Siberia, 18 乌苏里地区Ussuri region, 19 蒙古Mongolia, 20 帕米尔高原Plateau of Pamir, 21 中国东北Northeastern China, 22 中国西北Northwestern China, 23 中国青藏高原Qinghai-Xizang plateau of China, 24 中国西南Southwestern China, 25 中国华南Southern China, 26 中国中东部Centre-eastern China, 27 中国台湾Taiwan region of China, 28 朝鲜半岛Korea Peninsula, 29 日本Japan, 31 喜马拉雅地区Himalayan region, 32 印度与斯里兰卡Indian and Sri Lanka, 33 缅甸Myanmar, 34 中南半岛Indochina Peninsula, 35 菲律宾Philippines, 36 印度尼西亚Indonesia, 37 新几内亚New Guinea, 38 太平洋岛屿Islands of Pacific Ocean, 41 北非Northern Africa, 42 西非Western Africa, 43 中非Central Africa, 44 刚果河流域Congo river basin, 45 埃塞俄比亚地区Ethiopia region, 46 坦桑尼亚地区Tanzania region, 47 安哥拉地区Angola region, 48 南非South Africa, 49 马达加斯加Madagascar, 51 西澳大利亚Western Australia, 52 北澳大利亚Northern Territory, 53 南澳大利亚South Australia, 54 昆士兰Queensland, 55 新南威尔士New South Wales, 56 维多利亚Victoria, 57 塔斯马尼亚Tasmania, 58 新西兰New Zealand, 61 东加拿大Eastern Canada, 62 西加拿大Western Canada, 63 美国东部山地Mts. Eastern US, 64 美国中部平原Plain Central US, 65 美国中部丘陵Hills Central US, 66 美国西部山地Mts. Western US, 67 墨西哥Mexico, 68 中美地区Central America region, 69 加勒比海岛屿Caribbean Islands, 71 委内瑞拉Venezuela, 72 圭亚那高原Plateau Guyana, 73 安第斯山北段Northern Mt. Andes, 74 亚马孙平原Amazon Plain, 75 巴西高原Plateau Brazil, 76 玻利维亚Bolivia, 77 阿根廷Argentina, 78 安第斯山南段Southern Mt. Andes
01 北欧Northern Europe, 02 西欧Western Europe, 03 中欧Central Europe, 04 南欧Southern Europe, 05 东欧Eastern Europe, 06 俄罗斯欧洲部分European Russia, 11 中东Middle East, 12 沙特阿拉伯Saudi Arabia, 13 也门与阿曼Yemen and Oman, 14 伊朗高原Plateau of Iran, 15 中亚Central Asia, 16 西西伯利亚Western Siberia, 17 东西伯利亚Eastern Siberia, 18 乌苏里地区Ussuri region, 19 蒙古Mongolia, 20 帕米尔高原Plateau of Pamir, 21 中国东北Northeastern China, 22 中国西北Northwestern China, 23 中国青藏高原Qinghai-Xizang plateau of China, 24 中国西南Southwestern China, 25 中国华南Southern China, 26 中国中东部Centre-eastern China, 27 中国台湾Taiwan region of China, 28 朝鲜半岛Korea Peninsula, 29 日本Japan, 31 喜马拉雅地区Himalayan region, 32 印度与斯里兰卡Indian and Sri Lanka, 33 缅甸Myanmar, 34 中南半岛Indochina Peninsula, 35 菲律宾Philippines, 36 印度尼西亚Indonesia, 37 新几内亚New Guinea, 38 太平洋岛屿Islands of Pacific Ocean, 41 北非Northern Africa, 42 西非Western Africa, 43 中非Central Africa, 44 刚果河流域Congo river basin, 45 埃塞俄比亚地区Ethiopia region, 46 坦桑尼亚地区Tanzania region, 47 安哥拉地区Angola region, 48 南非South Africa, 49 马达加斯加Madagascar, 51 西澳大利亚Western Australia, 52 北澳大利亚Northern Territory, 53 南澳大利亚South Australia, 54 昆士兰Queensland, 55 新南威尔士New South Wales, 56 维多利亚Victoria, 57 塔斯马尼亚Tasmania, 58 新西兰New Zealand, 61 东加拿大Eastern Canada, 62 西加拿大Western Canada, 63 美国东部山地Mts. Eastern US, 64 美国中部平原Plain Central US, 65 美国中部丘陵Hills Central US, 66 美国西部山地Mts. Western US, 67 墨西哥Mexico, 68 中美地区Central America region, 69 加勒比海岛屿Caribbean Islands, 71 委内瑞拉Venezuela, 72 圭亚那高原Plateau Guyana, 73 安第斯山北段Northern Mt. Andes, 74 亚马孙平原Amazon Plain, 75 巴西高原Plateau Brazil, 76 玻利维亚Bolivia, 77 阿根廷Argentina, 78 安第斯山南段Southern Mt. Andes
Figure 1. BGUs of the world
图1. 世界陆地的基础地理单元
Records, BDR)将是定量分析的基础材料。各BGU的双子叶植物主要类群属数如表2。
2.4. 聚类分析方法
我们提出的相似性通用公式突破了二元比较的束缚。它的定义是:多个地区间的相似性系数是参加分析的各个地区的共有种类的平均数占总种类的比例 [16] :
式中,SIn是n个地理单元的相似性系数,Si,Hi和Ti分别是i地理单元的种类数、共有种类(common species)数、独有种类(unique species)数,且满足Hi = Si − Ti,Sn是n个地理单元的总种类数。计算时所需各个数值都可以很方便地从数据库的查询页面上获得。无论手工计算或计算机软件分析都非常方便快捷。
与SGF配套使用的多元相似性聚类分析法(MSCA)是任何组群的相似性系数都由参与分析的BGU 原始数据直接计算,不受先行计算的相似性系数的影响,也不受聚类顺序的限制。甚至可以先行计算67 个BGU 的总相似性系数。最后按相似性系数大小排列聚类图 [17] 。总相似性系数(General similarity coefficient, GSC)是传统分析方法所没有的概念和无法计算的指标。
3. 结果
世界20809属双子叶植物的聚类结果(图2)显示,67个BGU的总相似性系数为0.101,在0.410的相似性水平上,67个BGU聚成a-t共20个小单元群(Small unit crowd, SUC),在0.250的相似性水平上又聚成A-G共7个大单元群(Large unit crowd, LUC)。各群的组成单元都相邻相连,符合地理学原则;各群内的相似性程度大于群间的水平,符合统计学原则;各群的生态环境相对独立,符合生态学原则。各个大群、小群都有一定比例的特有属(表3),符合生物学原则。
对双子叶植物各主要类群的分析结果,都分别聚成7~8个大单元群和18~22个小单元群。各个聚类结果之间没有显著差异(表4),各群的组成基本一致。因此可以相信,图2的分析结果不带有偶然性,而是综合体现了各个类群的分布特征。
聚类结果与Cox建议的现行世界植物区划相比 [10] ,C、D、E三大群与印度–太平洋界、非洲界、澳洲界相同,主要不同有二:一是A、B、F三大群相当于把泛北极界分作三部分;二是G大群相当于新热带界划出中美地区给F大群。
聚类结果与华莱士的哺乳动物的区划相比 [50] ,D大群与非洲界相同外,主要不同有三:A、B两大群相当于把古北界分为东、西两部分;C、E两大群相当于把澳洲界的新几内亚岛、太平洋岛屿划归东洋界;F、G两大群相当于把新热带界的中美地区划归新北界。
聚类结果与世界昆虫区划 [24] [25] 相比,表现出高度的一致性,大、小群的数量完全相同,各大、小群的组成基本一致,群间的结构基本一致。所不同的是,总相似性系数及划分大、小单元群的相似性水平略高于昆虫,这是由于人们对植物比较关注,调查比较深入所致;个别单元的聚类位置有所移动,大群间只有25、31号地理单元互换位置。大群内也有个别地理单元在相邻小群之间移动,这些都不违背地理学原则。
因此,可以将聚类结果作为修订世界植物地理区划的定量依据,7个界依次命名为西古北界、东古北界、印度–太平洋界、非洲界、澳大利亚界、新北界和新热带界。
4. 讨论
本研究用MSCA方法首次较好地解析了世界各地双子叶植物的分布格局,比目前常用分析方法得到合理准确的聚类结果,又很接近现行的定性分析的世界植物地理区划方案。可以作为支持并局部修正
Table 2. The number of genus in every BGU of main groups of global Magnoliopsida plant
表2. 世界双子叶植物主要类群在各BGU的属数
BDR: Basic Distributional Records,基础分布记录;BBU: Basic Biological Unit,基础生物单元;ADT: Averagy Distributional Territory (BDR/BBU),平均分布域(基础分布记录/基础生物单元)。
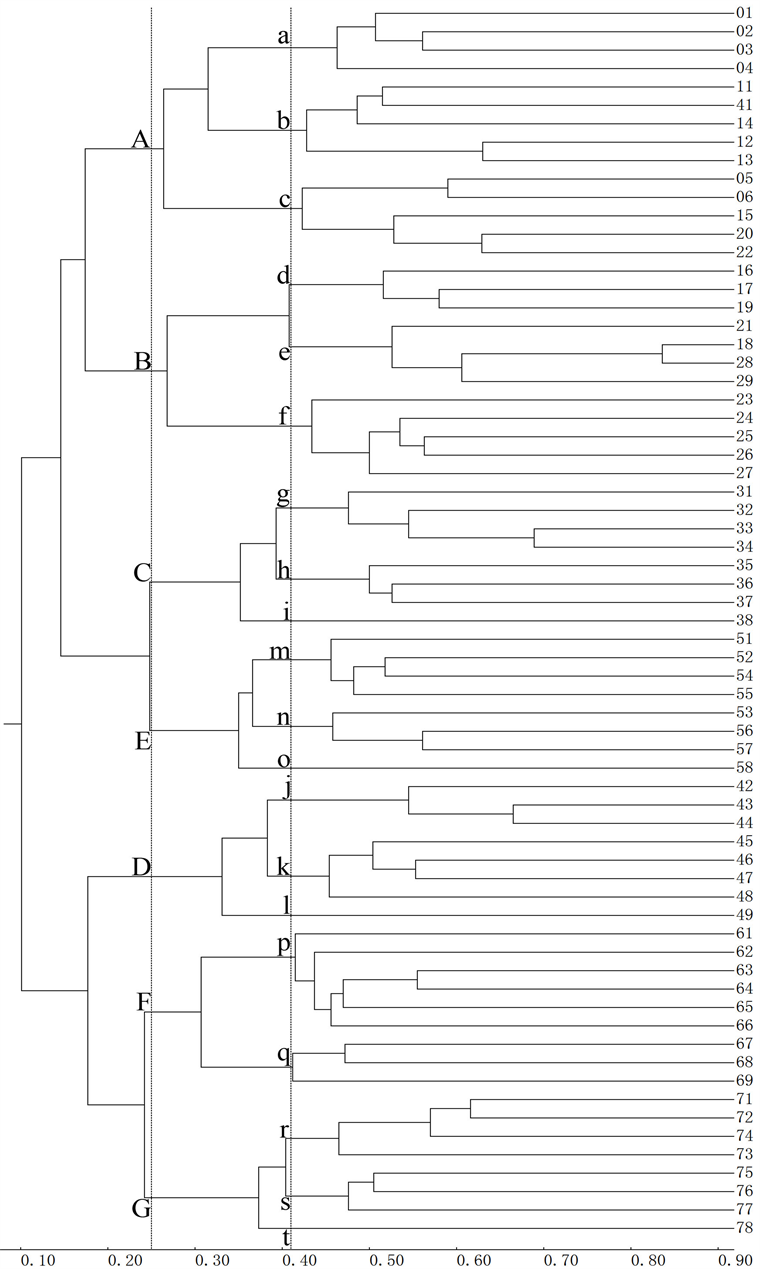
Figure 2. Dendrography of Magnoliopsida by MSCA
图2. 双子叶植物的MSCA法聚类图
Table 3. Magnoliopsida flora of every large and small unit groups
表3. 各单元群的双子叶植物区系
Note: LUC: Large unit crowds; SUC: Small unit crowds; BGU: Basic Geographical units.
Table 4. Analysis results of every main group of Magnoliopsida
表4. 双子叶植物各主要类群的分析结果
世界植物地理区划的定量依据。可以进一步补充单子叶植物、裸子植物以及苔藓植物等,以完善并确定建立定量的世界植物地理区划方案。
本分析结果与世界昆虫的分析结果极其相似,体现了植物与以植食性为主的昆虫之间的食物链关系。据此,是否可以推断,世界动物直接或间接地取食于植物,也可能存在相似的分布格局。微生物大都是分解植物残体的,是否也应该与植物有相似的分析结果。因此建议在对动物与微生物进行定量分析后,重新制定各大类生物的地理区划方案或总的世界生物地理区划系统。
致谢
我们感谢世界各地学者,如英国伦敦国王学院C. Barry Cox教授,德国格丁根大学Holger Kreft教授,美国克莱姆森大学John C. Morse教授,美国犹他大学Daniel R. Gustafsson教授,斯洛伐克科学院地理研究所Peter Vrsansky教授,法国医学院Jean-Claude Beaucournu教授,英国牛津大学Robert J. Whittaker教授,捷克兽医及制药大学Tomas Najer教授,法国巴黎大学Maram Caesar 教授,巴西圣保罗大学Michel P. Valim教授,美国加利福尼亚州州立大学Miklos D. F. Udvardy教授,德国格赖夫斯瓦尔德大学Nikki H.A. Dagamac教授,爱沙尼亚塔尔图大学Leho Tedersoo教授,美国新墨西哥大学Jennifer A. Rudgers 教授,德国约翰古登堡大学Janine Fröhlich-Nowoisky教授,美国加利福尼亚州立大学欧文分校Kathleen K. Treseder教授,瑞士洛桑大学Antoine Guisan教授,澳大利亚维多利亚博物馆Kevin C. Rowe教授,老挝国立大学Daosavanh Sanamxay教授,泰国宋卡王子大学Pipat Soisook教授,巴西戈亚斯联邦大学M. V. Cianciaruso教授,匈牙利自然历史博物馆Gabor Csorba教授,巴西帕拉伊巴联邦大学Anderson Feijo教授,墨西哥国立大学Tania Escalante博士,巴西国立癌症研究所Cibele R. Bonvicino教授,智利康塞普西翁大学Daniel González-Acuñad教授等,或赠送文献,或修饰文稿,或深入讨论,或提出建议。
基金项目
河南省重点实验室专项基金(112300413221)。
NOTES
*通讯作者。