1. 引言
古北短丝蜉(Siphlonurus palaearcticus)隶属节肢动物门(Arthropoda),昆虫纲(Insecta),蜉蝣目(Ephemeroptera)短丝蜉科(Siphlonuridae)短丝蜉属(Siphlonurus Eaton) [1] ,是一种分布范围广的净水性种类,在山区溪流中的底栖生物中具有代表作用。触角作为昆虫的重要感觉器官,其上着生有许多感受器 [2] [3] 。这些感受器调节着昆虫与外界物理、化学等各种环境因子的关系,在昆虫搜寻食物、躲避天敌和寻找配偶等行为中具有重要作用。
随着电子显微技术的发展,许多学者利用扫描电子显微镜对不同类型的昆虫触角感器进行了研究。1964年,Schneider对昆虫触角做了详细的描述:触角上分布很多由表皮特化形成的触角感器 [4] 。感器由毛原细胞、感觉神经细胞和一部分辅助细胞组成 [5] 。1970年Steinbrecht借助于光学和扫描电子显微镜进行了一项家蚕蛾触角形态学的详细研究,报道了毛形感器和锥形感器具有嗅觉功能 [6] 。70年代后期,以Zacharuk为代表的科学家将扫描电子显微镜(SEM)和透射电子显微镜(TEM)广泛应用到触角感器的超微结构研究。国内研究触角感器的起步时间较晚,70年代初,陈德明、杜家纬先后研制并使用了触角电位仪。1980年,尹文英首次报道了棉红铃虫触角感器的扫描电镜观察,随后杜家纬详细论述了昆虫触角感器的种类、形态及感受机理 [7] 。此后,我国对于触角感器方面的研究逐渐增多,但有关水生昆虫幼虫的触角超微结构观察方面的研究未见报道。由于水环境的独特性,在触角感器的种类、数量和分布上必然与陆生昆虫有一定的差别。
本研究采用扫描电子显微技术,对古北短丝蜉幼虫触角感器的超微结构进行了研究。揭示了古北短丝蜉幼虫感器的种类和分布规律,探讨了水生昆虫触角感器的独特性,为进一步探索水生昆虫对底栖环境的识别机制和行为调控提供理论依据。
2. 材料与方法
2.1. 供试虫源
本文所选古北短丝蜉幼虫为2014年10月下旬,在黑龙江省尚志市帽儿山地区阿什河流域(表1),水深0.4 m的净水缓流区采集的。在全长5 km的河段内随机选择2个样点用D形抄网(0.3 m宽,40目尼龙纱)捞取枯落物底泥,每个样点选择三个面积0.6 m × 1 m的小样方。在野外分拣蜉蝣幼虫样本,并立即用60%酒精溶液装瓶固定。实验室内,根据周上发发表的《中国大陆蜉蝣目分类研究》 [1] 进行形态学分类鉴定,挑选出古北短丝蜉幼虫样本备用。

Table 1. Geography coordinate of the sampling sites
表1. 采样点经纬度坐标
2.2. 样品的制备与观察
在采集瓶中随机选择20只健康完整的古北短丝蜉幼虫标本,置于培养皿中,倒入60%酒精溶液使其恰好没过虫体。在体视显微镜下,用超细镊子将昆虫的触角与头部分离,注意保持触角的完整性。将分离的触角依次用70%、80%、90%、100%的丙酮漂洗并脱水,每个梯度漂洗三次,每次15分钟。底栖昆虫由于长期生活在枯落物淤泥中,触角上粘有许多杂质,在进行电镜标本制作时可适当增加漂洗时间和次数。
将经过漂洗和梯度脱水的触角用毛刷转移到洁净的滤纸上,置于背阴处,使触角上的丙酮自然挥发。将晾干的昆虫触角用毛刷转移到电镜实验台的导电胶带上,置于离子溅射仪(SCD 005, USA)中喷金镀膜。将喷金镀膜后的标本试验台置于Quanta200 (FEI Inc, USA)扫描电子显微镜(SEM)中,在10~20 kV加速电压下观察并拍照。
2.3. 数据处理
实验图像使用Photoshop CS 7.0进行处理,数据测量使用Ruler电子尺,数据统计和表格分析使用Excel软件。文中所有数据均是10个以上测量数据的平均值。
目前触角感器的命名还没有一个统一的标准,本文沿用Schneider的分类系统,并参考马瑞燕、余海中、那杰的分类方法,对古北短丝蜉幼虫的触角感器进行分类和命名 [8] [9] [10] [11] [12] 。
3. 结果与分析
3.1. 触角的一般形态
古北短丝蜉幼虫的触角呈刚毛状(图1),向头部外侧上方弯曲,弯曲弧度较小。触角由三个部分组成,柄节、梗节和鞭节。柄节通过触角窝与头部相连,其上感器数量较少。
触角全长1.19~1.42 mm,平均长1.27 ± 0.9 mm。其中柄节最长,平均长度为156.7 ± 20.0 μm,中部直径98.9 ± 24.6 μm;梗节较短,全长为79.7 ± 13.7 μm,中部直径67.4 ± 14.9 μm;鞭节由10个圆筒形索节构成,第7索节最长,为150.4 μm,然后向两侧长度逐渐递减,第10索节特化成圆锥形的探索区,长度为58.3 μm,各索节中部直径由按1~10索节的顺序逐渐递减,平均宽度约44.6 μm (图2)。
触角表面密被鳞片,柄节鳞片大小不一,排列无明显规律,梗节和鞭节鳞片同为扁圆形且整齐有序。触角感器着生于鳞片之间的凹陷处,分布较稀疏。
3.2. 触角感器的种类、形态和分布
利用扫描电子显微镜对古北短丝蜉幼虫触角进行观察,共发现4种触角感器,分别为毛形感器(Sensilla trichoidea, ST)、耳形感器(Sensilla auricillica, SA)、钟形感器(Sensilla campaniformia, SC)和微孔感器(Sensilla microporous, SM)。

Figure 1. Full figures for outside A and inside B of antennal sensilla in Siphlonurus palaearcticus larva (A and B magnified 100 times)
图1. 古北短丝蜉幼虫触角外侧面A和内侧面B全图(A、B放大倍数均为100倍)
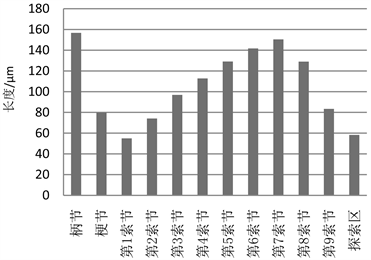
Figure 2. Statistics for each section length of Siphlonurus palaearcticus larva antenna (mean value)
图2. 古北短丝蜉幼虫触角各节长度(平均值)统计
3.2.1. 毛形感器
该感器为长条形刚毛状,从基部到顶端逐步变细。基部着生于表皮凹陷的感器窝内,尖端与触角末端同向。毛形感器是古北短丝蜉幼虫触角上较多的一种感器,触角各节上均有分布,尤其是探索区。根据其外形和长度可分为STI和STII两种类型,其中STI型触角长32.93 ± 4.63 μm,基部直径1.91 ± 0.06 μm,与触角表面呈60˚倾斜,在近端部1/3处弧形弯曲(图3A);STII感器长度为27.47 ± 6.94 μm,基部直径2.01 ± 0.09 μm,近基部突然弯曲,贴于触角表面,几乎与触角平行(图3B)。
3.2.2. 耳形感器
形似兔耳,下部宽扁,上部稍尖,端部及两侧向外卷曲成耳状(图4),表面有纵向纹理,基部着生于表皮凹陷的孔洞内。平均长度为11.24 ± 2.09 μm,基部直径2.08 ± 0.19 μm。分布在触角柄节、梗节和鞭节的1~8索节端部,平均每索节约有4个。
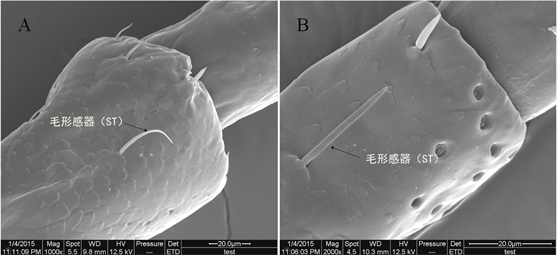
Figure 3. Sensilla trichoidea (A: magnified 1000 times, B: magnified 2000 times)
图3. 毛形感器ST (A:放大1000倍,B:放大2000倍)
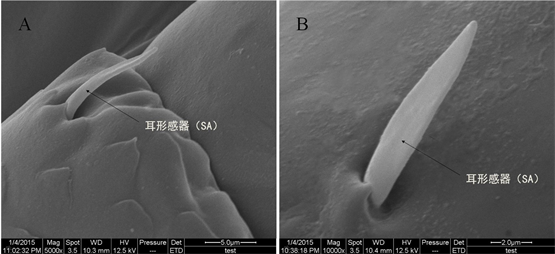
Figure 4. Sensilla auricillica (A: magnified 5000 times, B: magnified 10000 times)
图4. 耳形感器SA (A:放大5000倍,B:放大10000倍)
3.2.3. 钟形感器
形似纽扣,着生表皮凹陷的圆形小穴内,基部有一圈光滑的厚壁组织,中间有乳状突起(图5)。平均宽度为3.35 ± 0.26 μm,乳突直径1.28 ± 0.62 μm。密集分布在鞭节各索节相接处,靠近触角端部的下缘。平均每个索节上约有8~10个。

Figure 5. Sensilla campullacea (A: magnified 1000 times, B: magnified 3000 times)
图5. 钟形感器SC (A:放大1000倍,B:放大3000倍)
3.2.4. 微孔感器
极细小的孔状结构,中间有深陷孔内的小疣突(图6)。小孔直径为1.06 ± 0.07 μm,小疣突0.51 ± 0.04 μm。该感器数量极少,且仅分布在鞭节第三索节的外侧。

Figure 6. Sensilla microporous (A: magnified 1000 times, B: magnified 3000 times)
图6. 微孔感器SM (A:放大1000倍,B:放大3000倍)
4. 讨论
国内外对昆虫触角感器的形态结构和功能关系已有很多研究。大量资料表明毛形感器具有机械感受和化学感受的双重功能 [13] 。因此毛形感器在昆虫取食和生境选择中对环境的感知具有重要作用。古北短丝蜉幼虫触角上的毛形感器触角各节上均有分布,尤其是鞭节第十索节的探索区,推测其与探测物体形状、位置有关。另一方面,毛形感器较大的表面积使其容易接触水中的各种化学物质,从而实现嗅觉功能。本研究观察到古北短丝蜉幼虫触角上有两种毛形感器,根据形态结构与生理功能的关系可推测这两种毛形感器的具体功能。STI型毛形感器突出于触角表面,应该主要具备机械感知的作用。STII型毛形感器紧贴触角表面,且比表面积较大,容易接触水环境中的化学物质,可能具有化学感知的功能。
耳形感器突出于触角表面,广布于触角各节相接处的前沿。统计表明耳形感器在各小节的分布十分均匀,且相对数量较多,是古北短丝蜉幼虫触角上常见的触角感器。超微观察表明,该感器壁很薄,表面布满大量小孔,推测其具有嗅觉受体的作用 [14] 。古北短丝蜉幼虫触角上如此多的耳形感器,可能是为了适应溪流底部较复杂的化学环境。
钟形感器位于古北短丝蜉幼虫触角鞭节下缘,不利于接触物体表面。相对于其他触角感器,钟形感器的横截面积较大。推测其具有机械感知的功能,与感受表皮压力有关 [15] 。古北短丝蜉幼虫触角上的钟形感器数量最多,这可能是为了对水中复杂的暗流等进行判别。
微孔感器,也有文献描述为腔形感器,是一种深埋触角内部且数量极少的感受器。推测其具有感受温度和湿度变化的作用 [16] 。微孔感器在古北短丝蜉幼虫触角上并不常见,每条触角上只有1~2个,很可能是一种特殊的触角感器。
昆虫感器的形态、数量和分布等特点在昆虫进化及系统发生中具有重要意义,是昆虫经过长期自然选择的结果 [17] [18] 。研究昆虫触角感器的形态结构、生理功能及分布规律,对于理解昆虫与环境的协同进化具有指导性作用。而这种协同进化在宏观上又反映了水生态系统的稳定性。相关资料表明昆虫生活环境越简单,感器的类型和数量就越少。古北短丝蜉幼虫长期在净水区的浅层河床中生活,其生存的环境较为稳定,而砂石及枯落物又为其提供了躲避天敌的天然住所,所以触角感器种类明显不如陆生昆虫复杂且感器数量相对较少。据此推测其他水生昆虫的触角感器种类、数量和功能也相对简单,这与其生存的水环境有直接关系。
此外,昆虫的感受器并非仅分布在触角上,在虫体其他位置也有分布 [19] 。本文只对古北短丝蜉幼虫的触角感器进行了初步的形态观察,关于古北短丝蜉幼虫的其它体表感受器,各触角感器的具体作用以及古北短丝蜉成体触角感器分布在雌雄两性之间的具体差异还需要扩大样本数量,进行更加细致的分类研究。
5. 结论
结果表明,古北短丝蜉幼虫具有两种毛形感器、一种耳形感器、一种钟形感器和一种微孔感器。四种感器在触角上的分布规律并不相同:其中毛形感器分布最广,从触角柄节、梗节到鞭节第十索节的探索区均有分布;但感器数量最多的是钟形感器,在鞭节各索节相接处的端部内侧均有分布,且数量较多;耳形感器分布在触角柄节、梗节和鞭节的第1~8索节上,一般分布在各节端部,但中部也略有分布;微孔感器只分布在鞭节第3索节的中部,且数量极少,一般每条触角上只有1~2个。利用扫描电子显微镜对昆虫超微结构进行观察,是在微观层面上研究其结构与功能以及与环境因子的关系的一条有效途径。本文将这一途径引入对水生昆虫的研究,在古北短丝蜉幼虫触角感器超微结构观察的过程中,优化了实验流程,为进一步研究其他水生昆虫的超微结构奠定了基础。
致谢
感谢东北林业大学林学院孟昭军老师在昆虫触角采集和处理过程中给予的帮助和指导。
NOTES
*通讯作者。