1. 引言
恐惧是最常见的情绪之一,它在人类生存中起着至关重要的作用(Zsido, Deak et al., 2018)。根据注意的认知资源理论,认知系统会把认知资源分配到重要的刺激上。同时认知系统内还存在着一个评价环境重要性的系统,重要的刺激会优先进入到这个系统,获得进一步加工(彭聃龄,2012)。因此,占有认知资源比较多、优先加工的刺激会引起注意偏向。威胁信息的注意偏向一直以来都被广泛的讨论,并且大部分研究者都观察到个体对于威胁刺激存在优于中性刺激的注意偏向(牟柳&刘尚礼,2017)。从进化心理学角度,对威胁刺激进行快速反应在自然环境中极占优势,物种是否具备这种自适应功能会制约其进化与发展(Gernot, 2009)。
蛇、蜘蛛这类物种自古以来就对人类具有较大的生存压力,属于强进化相关威胁刺激的典型代表(Ohman, 2009; Hakamata et al., 2010)。Lobue以蛇、蜘蛛图片为刺激,发现被试搜索威胁图片的速度要快于搜索中性图片的速度,从而证明个体对于强进化相关威胁刺激具有注意偏向(LoBue & Matthews, 2014)。王福兴等人采用4~6岁没有经验的儿童作为被试,发现即使是对于蛇、蜘蛛没有经验的儿童,仍然存在对强进化相关威胁刺激的注意偏向,进一步证明人们对于强进化相关威胁刺激的注意偏向是先天的(王福兴,李文静,颜志强,段朝辉,&李卉,2015)。针对这一现象,Ohman等人认为,那些人类发展史中威胁生存的刺激能够快速的捕获人们的注意,使人们及时处理逃离危险。人类为了提高生存率,进化出了一个恐惧模型,专门处理与进化相关的威胁刺激(Ohman, Flykt, & Esteves, 2001)。但也有人提出个体对于蛇、蜘蛛的注意偏向可能是由刺激的恐惧程度引起的,而不是被试内在的恐惧模块加工的结果(Lipp & Waters, 2007)。如Zsido等人研究表明,一些现代的具有威胁性的物体也能引起同样的反应(Zsido, Bernath, Labadi, & Deak, 2018)。Fox等人在此基础上,采用与蛇、蜘蛛威胁程度相同的刀、枪作为弱进化相关威胁刺激,研究发现被试对于刀、枪的搜索反应时显著小于中性刺激,说明个体对于弱进化相关的威胁刺激也存在相似的注意偏向(Fox, Griggs, & Mouchlianitis, 2007),这在一定程度上反驳了恐惧模型理论。因为刀、枪是近现代才出现的,属于弱进化相关的威胁刺激,无法用恐惧模型进行解释。
大部分关于进化相关威胁刺激的研究集中于探究注意偏向,较少涉及对其记忆优势的研究。虽然已有研究发现人们对那些生存相关的刺激存在显著的记忆优势效应(Wells, Beevers, Robison, Ellis, 2010;王乾东,李庆功,陈凯凯,&傅根跃,2015),如有研究者以蛇和乌龟为对象,研究人们对于强进化相关威胁信息和中性信息的记忆优势,得出被试对于乌龟的再认正确率要显著低于蛇(柴江霞,肖玉珠,姚伟莉,&赵笑梅,2015),说明人们对于强进化相关威胁刺激具有显著的记忆优势。负性情绪面孔也常被研究者用作威胁性刺激,Yamagishi等人通过拍摄囚徒困境博弈中真实的合作者和欺骗者面孔照片进行再认实验。结果发现,欺骗者的面孔同样比合作者面孔具有更高的再认准确率(Yamagishi, Tanida, Mashima, Shimoma, & Kanazawa, 2003)。但这类实验目前的研究较少,而且方法比较单一,到现在也没有一个统一的结论。
2. 问题提出
本模板仅针对采用A4纸型的论文版式。请务必确保您的论文采用A4幅面(21厘米 × 28.5厘米)进行排版。上述研究均证明了个体对于进化相关威胁刺激存在注意偏向,但该注意偏向是否与人类的进化相关还存在争论。并且,由于这类实验的研究范式比较单一,均采用视觉搜索范式,所以不能很好的说明被试对于进化相关威胁刺激的注意偏向具体表现在哪些成分上,是注意解离困难还是加速,所以本实验以点探测范式(Koster, Crombez, Verschuere, Vanvolsem, & De Houwer, 2007;毋嫘,林冰心,蒋娜,&赵亚萍,2016)为任务,并增加中性刺激对作为基线,将被试在中性与一致性条件下的任务反应时进行对比,并探究强和弱进化相关威胁刺激的注意成分是否存在差异。
其次,有研究证明相较于其他刺激,强进化威胁刺激存在记忆优势(Wells, Beevers, Robison, & Ellis, 2010;柴江霞,肖玉珠,姚伟莉,&赵笑梅,2015),但对于弱进化相关威胁刺激的研究很少。所以,本研究采用代表强进化相关的刺激(蛇,蜘蛛)与弱进化相关的刺激(枪,刀)等威胁性图片研究个体对于两种类型的威胁性刺激是否存在记忆优势,并且探究个体对于这两种威胁性刺激的记忆优势是不是存在差异。
除此之外,有研究(Ohman, Flykt, & Esteves, 2001)证明个体对于威胁性刺激的注意偏向是相对自动的、无需努力的过程,威胁信息的自动化加工使个体能够迅速做出逃跑或攻击的决定,从而提高生存概率(Boyer & Bergstrom, 2011)。但是关于认知负荷和威胁刺激类型之间的相互作用还不是很清楚。所以,本研究在第一个实验的基础上加上一个认知负荷任务来探讨认知负荷对于个体注意偏向的影响以及不同认知负荷条件下强或弱进化相关刺激的注意偏向是不是存在差异。
3. 实验1点探测任务下对于进化相关威胁刺激的注意偏向
3.1. 实验目的
本实验运用点探测范式(Koster, Crombez, Verschuere, Vanvolsem, & De Houwer, 2007;毋嫘,林冰心,蒋娜,&赵亚萍,2016)来研究个体对于进化相关威胁性刺激的注意偏向,以及探讨个体对于强和弱进化相关威胁刺激的注意偏向有无差别。并且在实验中加入中性–中性图片对来探究个体对于强和弱进化相关威胁刺激的注意偏向究竟表现在哪个成分上,是注意定向加速还是解除困难。分别将一致和不一致性条件下探测任务的反应时和中性条件进行比较,当中性条件下的注意反应时大于一致性条件为注意定向加速;当中性条件下的注意反应时小于不一致条件下为解除困难。除此之外,实验最后通过一个再认任务来了解个体是否存在对于强和弱进化相关威胁刺激的记忆优势,以及两者之间是否有显著的差异。
3.2. 实验假设
1) 个体对于强和弱进化相关威胁性刺激均存在注意偏向,并且个体对于强进化相关威胁性刺激具有显著优于弱进化相关威胁性刺激的注意偏向;
2) 在SOA为500 ms的条件下,个体对强进化相关威胁刺激和弱进化相关威胁刺激的注意偏向表现为注意定向加速和解除困难;
3) 相较于中性刺激,个体对于强、弱进化相关威胁刺激都具有较高的再认正确率,并且强进化相关威胁性刺激的再认正确率高于弱进化相关威胁性刺激。
3.3. 方法及程序
3.3.1. 被试
实验选用年龄在20~22岁的大学生被试32名,其中男生13人,女生19人,此前都没有参加过这类实验,视力行为能力均良好。
3.3.2. 实验材料
刺激材料是参考相关文献(Brown, El-Deredy, & Blanchette, 2010; Ohman, Flykt, & Esteve, 2001)选自于互联网,一共230张。其中包括50张强进化相关威胁图片(蛇、蜘蛛),50张弱进化相关威胁图片(刀、枪),130张中性图片(花、水果、盆、桶)。将这些图片交由20位非正式实验中的被试进行威胁程度的7点评分(文少司,2015) (1代表完全没有威胁,7代表非常大的威胁)。最终得出强进化相关威胁刺激和弱进化相关威胁刺激的威胁程度差异不显著;强进化相关威胁刺激的威胁程度明显高于中性刺激;弱进化相关威胁刺激的威胁程度明显高于中性刺激。
实验1随机的从上述图片中选取130张图片作为材料,包括代表强进化相关威胁刺激的图片30张;代表弱进化相关威胁刺激的图片30张;以及70张图片代表中性刺激(见附录)。其中30张作为再认任务的材料,强、弱进化相关和中性刺激图片各10张。剩余100张图片组成三种类型的图片对,其中20张强进化相关威胁图片和20张中性图片组成强进化–中性图片对,20张弱进化相关威胁图片和20张中性图片组成弱进化–中性图片对,还有20张中性图片组成中性–中性图片对。
3.3.3. 实验程序
整个实验分为点探测任务和再认任务,点探测任务一共55个试次,其中5个试次为练习任务,50个是正式实验任务,50个试次呈现的图片对分为20个强进化–中性刺激对、20个弱进化–中性刺激对和10个中性–中性刺激对。在前两个类型的刺激对中,强进化、弱进化和中性刺激出现在左右两侧的概率相等,并且探测点出现在左右两侧概率也一致。再认任务一共60个试次,其中30个试次中呈现的图片是之前实验中呈现过的,另外30个试次中的图片在先前的实验中没有呈现过。实验为2 (刺激类型) × 2 (试次类型)的被试内设计,实验流程如图1所示。
3.4. 结果及分析
实验共收集了32名被试的数据,由于实验的错误率(0.8%)过低,所以只对实验的反应时进行分析。并且删除3个极端反应时和三个标准差之外的反应时数据。
实验以被试探测任务的平均反应时作为因变量,刺激类型(强进化相关威胁刺激和弱进化相关威胁刺激)和实验试次类型(一致条件和不一致条件)为自变量进行2 × 2的重复测量方差分析,分析结果如图2所示。
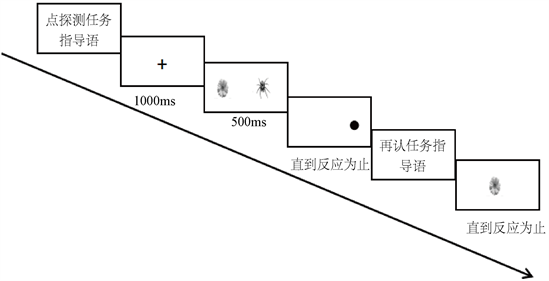
Figure 1. Experimental flow chart of point detection under no cognitive load
图1. 无认知负荷下点探测实验流程图
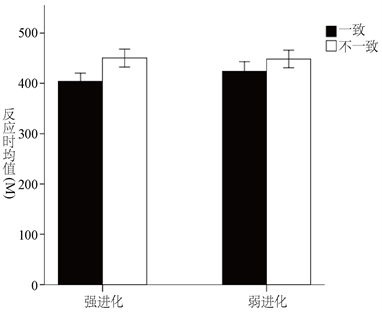
Figure 2. Time difference in attention response to evolutionary-related threat stimuli without cognitive load
图2. 无认知负荷时进化相关威胁刺激的注意反应时差异图
结果表明,刺激类型的主效应不显著,F(1,31) = 2.621,p = 0.116,被试对于强进化相关威胁刺激和弱进化相关威胁刺激的反应时差异不显著;试次类型的主效应显著,F(1,31) = 19.897,p < 0.0005,η2 = 0.391,表明被试在不一致条件下的探测任务反应时显著大于一致条件下的探测任务反应时;刺激类型和试次类型的交互作用不显著,F(1,31) = 2.804,p = 0.104。
由于实验结果只能表明被试对于进化相关威胁刺激存在注意偏向,而不能区分具体表现在注意的哪个成分上。为了区分这一点,本实验参照相关文献增加了一个中性–中性刺激对,用来区分被试对于强和弱进化相关威胁刺激的注意偏向表现为注意定向加速或解除困难还是两者均有。将不同条件下的进化相关威胁刺激的反应时与中性条件下的反应时进行俩俩相关t检验,结果表明,一致性条件下的强进化相关威胁刺激的注意反应时明显小于中性条件下的反应时,反应时之差为−21.744,被试表现为对于强进化相关威胁刺激的注意定向加速,t(31) = −2.086,p = 0.045,d = 0.369。而一致性条件下的弱进化相关威胁刺激的注意反应时与中性条件下的反应时差异不显著,反应时之差为−2.219,说明不存在注意定向加速,t(31) = −0.228,p = 0.821,d = 0.041。不一致性条件下的强进化相关威胁刺激的注意反应时显著大于中性条件下的反应时,反应时之差为24.397,被试表现为对于强进化相关威胁刺激的注意解除困难,t(31) = 2.291,p = 0.029,d = 0.405。不一致性条件下的弱进化相关威胁刺激的注意反应时显著大于中性条件下的反应时,反应时之差为22.513,被试表现为对于弱进化相关威胁刺激的注意解除困难,t(31) = 2.080,p = 0.045,d = 0.368。
实验对于三种类型的刺激的再认正确率进行单因素重复测量方差分析发现,如图3所示,进化相关威胁刺激再认正确率差异显著,F(2,62) = 10.962,p < 0.0005,η2 = 0.261。
 ***p < 0.001。
***p < 0.001。
Figure 3. The difference in recognition accuracy of evolutionarily related threat stimuli
图3. 进化相关威胁刺激的再认正确率差异图
由于三种类型图片的再认正确率差异显著,为了了解究竟是哪两者之间存在差异,分别对强和弱进化相关威胁刺激及中性刺激进行俩俩的相关样本t检验,如图3所示,结果表明强进化相关威胁性刺激的再认正确率显著低于弱进化相关威胁性刺激,t(31) = −3.597,p = 0.001,d = 0.636。弱进化相关威胁刺激与中性刺激的再认正确率差异显著,t(31) = 4.126,p = 0.000,d = 0.729,即弱进化相关威胁刺激的再认正确率显著高于中性刺激。强进化相关威胁刺激与中性刺激的再认正确率差异不显著,t(31) = 0.430,p = 0.670,d = 0.076。
4. 实验2认知负荷条件下对于进化相关威胁刺激的注意偏向
4.1. 实验目的
目前,对于进化相关威胁刺激的注意偏向是否会受到认知负荷的影响还存在分歧,有研究者认为对于进化相关威胁刺激的注意偏向需要认知资源的参与,当认知资源不足时,会降低被试对于进化相关威胁刺激的注意加工(文少司,2015)。也有研究者认为个体对于进化相关威胁刺激的注意偏向是相对自动化的过程,不受认知资源的干扰(Goodwin, Eagleson, Mathews, Yiend, & Hirsch, 2017; Ohman, Flykt, & Esteve, 2001)。实验2在实验1的基础上,加入认知负荷任务,来进一步研究认知负荷是否会降低个体对于强和弱进化相关威胁刺激的注意偏向;以及探讨在认知负荷条件下,强和弱进化相关威胁刺激的注意偏向是否存在差异,并且两者的注意偏向成分是否一致。
4.2. 实验假设
1) 认知负荷会降低被试对于强和弱进化相关威胁刺激的注意偏向,被试在认知负荷条件下的注意反应时会明显高于无认知负荷条件下的注意反应时;
2) 认知负荷条件下,强和弱进化相关威胁刺激的注意偏向存在显著差异,两种类型刺激的注意偏向均表现为注意解除困难。
4.3. 方法及程序
4.3.1. 被试
采用的被试为上一个实验中的32名大学生被试。
4.3.2. 实验材料
刺激材料一共100张图片,所有图片在实验1中已经评定过。包括代表强进化相关威胁刺激的图片(蛇、蜘蛛)和代表弱进化相关威胁刺激的图片(刀、枪)各20张;代表中性刺激的图片(花、水果、盆、桶) 60张。这些图片参照实验1组成三种匹配的图片对,其中强进化–中性图片对和弱进化–中性图片对各20对,还有10对中性–中性图片对。同样运用2345看图王软件对图片统一做了灰度处理,所有图片均没有背景并且大小一致。最后,随机生成50个由6个数字组成的字符串如“683924”,以及50个带星号的字符串如“***9**”。
4.3.3. 实验程序
整个实验一共55个试次,其中5个试次为练习任务,50个是正式实验任务,50个试次分为20个强进化–中性刺激对、20个弱进化–中性刺激对和10个中性–中性刺激对试次。强进化、弱进化和中性刺激以及探测点出现在左右两侧的几率相同。实验为2 (刺激类型:强进化相关威胁刺激和弱进化相关威胁刺激) × 2 (负荷类型:负荷和无负荷)的被试内设计,因变量是探测任务的反应时和威胁偏向指数,实验流程如图4所示。
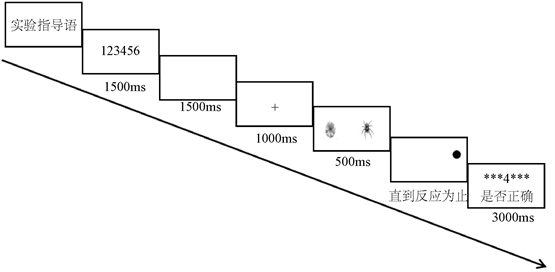
Figure 4. Experimental flow chart of point detection under cognitive load
图4. 认知负荷下点探测实验流程图
4.4. 结果分析
将实验数据像实验1一样处理,一共删除反应错误的数据(0.3%)和反应时 < 200 ms或者>1500 ms的数据11个。
以被试的平均反应时作为因变量,刺激类型(强进化相关威胁刺激和弱进化相关威胁刺激)和实验试次类型(一致性条件和不一致条件)为自变量进行2 × 2的重复测量方差分析,分析结果如图5所示。
结果表明,刺激类型的主效应不显著,F(1,31) = 0.230,p = 0.635,即强进化相关威胁刺激和弱进化相关威胁刺激的注意反应时差别不显著;试次类型的主效应显著,F(1,31) = 19.051,p < 0.0005,η2 = 0.381,即在不一致性条件下被试对进化相关威胁刺激的注意反应时显著慢于一致条件下的反应时;刺激类型和试次类型的交互作用不显著,F(1,31) = 1.243,p = 0.274。

Figure 5. Time differences in attention response to evolutionary-related threat stimuli under cognitive load
图5. 认知负荷下进化相关威胁刺激的注意反应时差异图
同样为了区分注意偏向的成分,增加了一个中性–中性刺激对,用来研究被试对于强和弱进化相关威胁刺激的注意偏向表现在哪个注意成分上。将被试在不同试次条件下与中性条件下对进化相关威胁刺激的注意反应时进行俩俩相关样本t检验,结果显示,一致性条件下的强进化相关威胁刺激的注意反应时与中性条件下的反应时没有明显差别,反应时之差为−9.772,被试没有表现出对于强进化相关威胁刺激的注意定向加速,t(31) = −0.927,p = 0.361,d = 0.164。一致性条件下的弱进化相关威胁刺激的注意反应时与中性条件下的反应时不存在显著差异,反应时之差为−6.956,注意定向加速不显著,t(31) = −0.595,p = 0.556,d = 0.105。不一致性条件下的强进化相关威胁刺激的注意反应时与中性条件下的反应时差异显著,反应时之差为33.719,被试表现为对于强进化相关威胁刺激的注意解除困难,t(31) = 3.167,p = 0.003, d = 0.560。不一致性条件下的弱进化相关威胁刺激的注意反应时与中性条件下的反应时差异显著,反应时之差为22.606,被试表现为对于弱进化相关威胁刺激的注意解除困难,t(31) = 2.070,p = 0.047,d = 0.366。
本实验为了探究认知负荷是否会影响被试对于进化相关威胁刺激的注意偏向,先分别计算不同类型刺激的威胁偏向指数,该指数是通过计算每个被试在中性线索下的反应时(即探测点出现在中性线索一侧)减去威胁线索下的反应时(即探测点出现在威胁线索一侧),如果该指数大于0,则表明被试的注意偏向于进化相关威胁刺激,如果该指数小于0,则表明被试的注意偏向于中性刺激。
以刺激类型(强和弱进化相关威胁刺激)和负荷类型(无负荷和负荷)为自变量,威胁偏向指数为因变量进行2 × 2的重复测量方差分析,分析结果如图6所示。
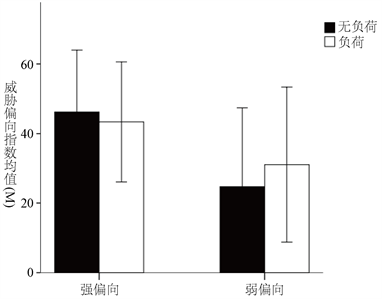
Figure 6. Threat bias index difference graph of different types of images
图6. 不同类型图片的威胁偏向指数差异图
结果表明,刺激类型的主效应不显著,F(1,31) = 3.754,p = 0.062,即强进化相关威胁刺激和弱进化相关威胁刺激的威胁偏向指数差异不显著;负荷类型的主效应不显著,F(1,31) = 0.010,p = 0.919,即认知负荷和无认知负荷条件下的威胁偏向指数没有明显差别;刺激类型和负荷类型的交互作用不显著,F(1,31) = 0.221,p = 0.642。因此,这些结果并没有证明认知负荷会对被试的注意偏向产生影响。
5. 讨论
已有不少研究证明个体对于进化相关威胁刺激存在注意偏向,Ohman等人从进化心理学的角度提出了恐惧模型的理论解释这一现象,认为个体对于进化相关威胁刺激的注意偏向是为了提高生存率发展而来的,是相对自动化的过程(Ohman, Flykt, & Esteve, 2001)。也有研究者对此持反对意见,认为根本不存在恐惧模型,并用枪等代表弱进化相关威胁刺激的研究得出这类威胁性刺激也能引起个体的注意偏向(Fox, Griggs, & Mouchlianitis, 2007),但枪是现代才出现的威胁人类生存的刺激,所以研究结果在一定程度上挑战了恐惧模型理论。并且,对于进化相关威胁刺激的研究大多采用视觉搜索范式,研究范式比较单一。因此,对于进化相关威胁刺激的注意偏向成分机制还不是很清楚。另外,以往有研究从进化心理学的角度出发证明了个体对那些与生命安全有关的信息具有较好的记忆优势(Nairne, Pandeirada, & Thompson, 2008),多数研究专注于对进化相关威胁刺激的注意偏向研究,本实验在此基础上研究不同类型的进化相关威胁刺激是否存在较好的记忆优势,并且探讨强和弱进化相关威胁刺激的记忆效果是不是存在差别。
为了探究以上问题,本实验设计了2个实验,实验1主要采用代表强进化相关的威胁刺激“蛇、蜘蛛”,代表弱进化相关的威胁刺激“刀、枪”,以及代表非威胁性刺激“花、水果、盆、桶”来研究个体是否对于进化相关威胁刺激存在注意偏向,并且表明具体体现在注意偏向的哪个成分上以及强进化相关威胁刺激和弱进化相关威胁刺激的注意偏向是否存在差异。最后用简单的再认任务来探究人们对于进化相关威胁刺激的记忆优势效应。结果与先前的研究结论一致(Brown, El-Deredy, & Blanchette, 2010),被试存在对强和弱进化相关威胁刺激的注意偏向,说明相较于中性刺激,被试更愿意把注意分配到进化相关威胁性刺激上,但强和弱进化相关威胁性刺激的注意偏向没有表现出明显的差异,这一结果不能证明恐惧模型理论,说明被试对威胁刺激的注意偏向与人类进化过程无关,结果无法用进化心理学的理论进行解释。产生该结果有可能是因为本实验采用的研究范式不是很适合用来研究强和弱进化相关威胁刺激的差异,单纯的反应时无法体现出两者注意偏向的区别,或者,强和弱进化相关威胁刺激注意偏向本身就不存在差别,也不存在加工强进化相关威胁刺激的恐惧模型。
再者,通过比较三种类型的威胁刺激再认正确率得出,强进化相关威胁刺激的记忆优势不明显,具体表现为强进化相关威胁刺激的再认正确率和非威胁刺激的差异不明显,这与以往的研究结果不一致(柴江霞,肖玉珠,姚伟莉,&赵笑梅,2015),这有可能是因为本实验的再认任务事先并没有告知被试而导致刺激没有得到很好的记忆加工。而弱进化相关威胁刺激表现出明显的记忆优势,弱进化相关威胁刺激的再认正确率显著高于非威胁刺激和强进化相关威胁刺激,说明弱进化相关威胁刺激得到了更多的注意加工和记忆,可能是相较于以前,现代的人们较少受到蛇、蜘蛛等强进化相关刺激的威胁,而更多的接触枪、刀等刺激,对于现代人类而言,弱进化相关威胁刺激更多的对生存造成威胁,为了提高生存率,人们更倾向于注意及加工这类刺激以便尽快做出反应,所以这类刺激得到更好的加工记忆,表现出更好的记忆优势。
为了进一步探究认知资源对注意偏向的影响,实验2在实验1的基础上加入了一个认知负荷任务,研究得出认知负载条件下被试仍然存在对进化相关威胁刺激的注意偏向,但认知负载条件下,无论是强进化相关威胁刺激还是弱进化相关威胁刺激在不一致条件下均表现为注意解除困难,这一点与以往关于威胁性刺激的研究结果相同(Goodwin, Eagleson, Mathews, Yiend, & Hirsch, 2017)。被试对两种类型的刺激均没有表现出注意定向加速,这与无认知负荷条件下被试对强进化相关威胁刺激存在定向加速有所不同,可能是因为点探测任务前的认知负荷任务消耗了被试的一部分认知资源,使得被试的注意无法快速的转移到威胁性刺激上去。最后,通过计算威胁偏向指数来代表被试偏向强和弱进化相关威胁刺激的程度,比较有无认知负荷条件下的威胁偏向指数得出,被试在认知负荷条件下对进化相关威胁刺激的注意偏向与无认知负荷条件下的注意偏向不存在显著差异,表明被试对于进化相关威胁性刺激的注意偏向是无意识的,很少受到认知资源的限制。这与Goodwin运用词汇代表威胁性刺激得出的实验结果一致(Goodwin, Eagleson, Mathews, Yiend, & Hirsch, 2017)。但也有研究者运用认知负载范式得出被试在认知资源匮乏的情况下对进化相关威胁刺激的注意偏向与无认知负载条件下的注意偏向存在显著差异,并且被试只存在对强进化相关威胁刺激的注意偏向,即认知资源会影响个体对进化相关威胁刺激的注意偏向,当认知资源严重不足时,被试不存在对弱进化相关威胁刺激的注意偏向(文少司,2015)。这表明了本实验没有观察到在不同负荷条件下被试的注意偏向的差异可能是由于认知负荷任务不足以占用足够多的认知资源导致的。
总体而言,本研究得出,被试对于强和弱进化相关威胁刺激存在相似的注意偏向,两者并没有表现差异,这一结果没能支持恐惧模型理论,与Brown等人得出的研究结果一致(Brown, El-Deredy, & Blanchette, 2010)。但也有研究者运用认知负载范式得出了不同的结果,提出强进化相关威胁刺激和弱进化相关威胁刺激在注意加工上是存在显著差异的(文少司,2015),从另一个角度说明了本实验没有表现出差异可能是由实验范式引起的。另外,额外的认知任务并没有显著降低或增加被试对于进化相关威胁刺激的注意反应时,表明被试对于进化相关威胁刺激的注意是不需要意志努力的过程,较少受到意志的控制。即使是在认知资源相对较少的条件下,被试依然能对进化相关威胁刺激进行加工。并且在有无认知负荷条件下均观察到了对进化相关威胁刺激的注意解除困难,这与一些研究者(Amir, Elias, Klumpp, & Przeworski, 2003)提出的刺激呈现500 ms以后不易观察到注意定向加速,容易观察到注意解除困难的结论一致。
6. 不足与展望
本研究通过2个实验探讨了个体对于进化相关威胁刺激的注意偏向和成分,以及认知负荷对于被试注意偏向的影响,得出了一些重要的结果,但仍然存在一些不足。
首先,本研究被试数量比较少,而且年龄段比较集中,另外本研究也没有从性别的角度探究注意偏向的差异,未来可以从年龄和性别上来分析个体对于进化相关威胁刺激是否存在差异。
其次,本研究只采用了图片作为材料代表进化相关威胁刺激,图片呈现的是刺激静态的样子,没有提供早期人类搜索蛇或蜘蛛时的必要环境信息。如果不提供必要的环境信息,现有的注意偏向结果也可能解释为新异刺激导致的效应(Peira, Golkar, Öhman, Anders, & Wiens, 2012)而日常生活中人们面对的更多的是动态的刺激,实验结果不能很好的说明在日常生活中人们是否依然对于这类刺激具有相同的注意偏向,在真实的视觉搜索中,物体远近及角度变化带来的动态信息对理解视觉搜索加工的认知机制很重要,而威胁性刺激呈现的情境信息(比如远古人类面对的丛林草地环境)对检验已有研究的生态效度同样重要。显然,无论是环境信息还是光流信息(袁小钧等,2018)。将来可以在改进实验材料,比如增加词汇、视频或者在接触真实的进化相关威胁刺激的基础上进行研究。
再次,本实验仅仅采用反应时和正确率作为判断标准,可能并不能很好的反应个体对于进化相关威胁刺激的注意偏向,并且,点探测范式是否是最适合用来研究注意偏向也还存在疑问,可能结合不同研究范式进行比较能得出更加准确的结果。另外,本实验还有一个局限是,我们不能确定所施加的负荷任务是否真正占用了足够的认知资源,虽然这里使用的负荷任务是以往研究中发现的占用大量的注意控制资源的典型负荷材料。所以,在反应时的基础上加上眼动和脑电等数据能够更加清晰的体现被试对于进化相关威胁刺激的注意偏向及成分机制。
最后对未来研究有两点展望:
1) 本研究刺激呈现时间固定,此间下的注意偏向前Mogg和Bradley (2006)使用视觉搜索范式研究了不同水平威胁刺激在不同暴露时。研究结果发现,当呈现时间为200 ms时,高水平威胁组的注意偏向比低水平组要大。随着暴露时间的增加,高水平组注意偏向降低。在更长暴露时间时,这种注意偏向差异则不再显著。这一研究表明,威胁性刺激对注意的捕获效应主要发生在早期的快速视觉信息加工阶段。在未来的研究中,可以考虑引入“刺激呈现时间”这一变量。
2) 在那些焦虑的人身上,不自觉地停留在环境的消极方面可能是频繁和过度的。研究结果明确支持了这样一个观点,即当焦虑存在时,威胁姿势会吸引注意力,而且是以一种无法控制的方式(Azarian, Esser, & Peterson, 2016)。未来的研究可以考虑与精神障碍结合研究,产生更多的现实意义。
7. 结论
(1) 人们普遍存在对于进化相关威胁刺激的注意偏向,该类刺激更能捕获人们的注意,但强和弱进化相关威胁刺激的注意偏向没有表现出差异;
(2) 在SOA为500 ms的条件下,被试对于强进化相关威胁刺激和弱进化相关威胁刺激的注意偏向均是由注意解除困难引起的;
(3) 被试存在对于弱进化相关威胁刺激的记忆优势,并且对于弱进化相关威胁刺激的记忆效果显著优于强进化相关威胁刺激;
(4) 个体对于进化相关威胁刺激的注意加工仅需要很少的认知资源,并且强和弱进化相关威胁刺激的注意加工耗费了相同程度的资源。
致谢
首先,非常感谢我的导师于欣教授以及二导周强副教授,本论文是在他们的悉心指导下完成的。作为我的导师,他们在我的整个论文选题设计到撰写论文完成都给予了很重要的指导意见,帮助我顺利完成整个实验,使我获益匪浅。其次,感谢王青同学对我在编程及数据处理方面的帮助和支持,尤其是在我处理数据遇到困难的时候细心的给我讲解。最后,感谢所有参加并认真完成实验的被试和所有为我的实验顺利完成提供见解的朋友们。
附录


NOTES
*通讯作者。